В.А. Бердников Эволюция и прогресс
АКАДЕМИЯ НАУК СССР СИБИРСКОЕ ОТДЕЛЕНИЕ Утверждено к печати Институтом цитологии и генетики СО АН СССРОтветственный редактор доктор биологических наук С.Н. Родин
Рецензенты доктора биологических наук А.С. Графодатский, Н.Д. Тарасенко
Введение
Для чего они? Откуда? Оправдать ли их умом? Но прекрасных тварей груда спит, разбросана кругом. И уйдет мудрец, задумчив, и живет как нелюдим, и природа, вмиг наскучив, как тюрьма стоит над ним.«От амебы до человека» — таким обычно представляется нам путь, пройденный эволюцией. Ну, а в старину, когда главным «инструментом» получения знаний о мире оставался невооруженный глаз наблюдателя, было модным противопоставление человека — «венца всего живущего» — «презренному червю, копошащемуся в прахе». Хотелось бы спросить, за что же червь был удостоен презрения? Вообще, почему слово «червь» стало синонимом слова «ничтожество»? Вероятно, за простоту строения и нетребовательность. Вот пчелу всегда уважали, и дело не в том, что она полезна (курица тоже полезна), но прежде всего в том, что она трудолюбива и отличается очень сложным, прямо-таки разумным поведением. Впрочем, высокая степень совершенства приписывалась и другим насекомым. Так, живший во II веке Лукиан написал «Похвалу мухе», хотя уже за тысячу лет до него оценил достоинство этого назойливого членистоногого Ромер. В «Илиаде» богиня Афина, вдохновляя царя Менелая к бою, укрепила его тело, а…Н.А. Заболоцкий
Сердце ж наполнила смелостью мухи, которая мужем,
Сколько бы крат ни была, дерзновенная, согнана с тела,
Мечется вновь уязвить, человеческой жадная крови,—
Смелость такая Атриду наполнила мрачное сердце.
Идея прогресса
Глава 1. Теории прогрессивной эволюции
Кто знает, что дух человека возносится ввысь, А дух скота — тот вниз уходит в землю?Бытовой антропоцентризм Человек может оставаться равнодушным ко многому, только не к собственной персоне. В себе его интересует буквально все: внешность, психика, умственные способности и особенно происхождение. Черты родителей и более отдаленных предков принято отыскивать и ценить. У всех народов существует представление о благородстве своих предков, иначе говоря, в той или иной форме существует их культ. Иногда, к большому сожалению, этот культ принимает форму национализма, и представители разных народов готовы на все, лишь бы доказать превосходство пращуров, а тем самым и свою исключительность. Забавно, что заложенное природой стремление к самовосхвалению заставляет нас выделить свой вид из остального животного мира и поставить его на высшую ступень. Согласитесь, что само название Homo sapiens, т. е. человек разумный, уже «звучит гордо». Умиляясь своим видовым особенностям, мы нередко вообще склонны отрицать наше родство с другими животными, ведя происхождение от какого-нибудь божества или, как теперь модно, от пришельцев из иных (внеземных) миров. Очевидное сходство с животными нередко даже задевает нашу гордость. Так, в книге Ч. Дарвина (1809–1882) «Происхождение человека и половой отбор» [1871] приводится одно объяснение, почему некоторые африканские племена выбивают себе клыки и красят зубы в синий цвет. Оказывается, они не хотят быть похожими на собак. Однако под напором неопровержимых свидетельств, с трудом признав свое родство с другими животными и согласившись попасть в одну группу с обезьянами, мы дали ей весьма нескромное название «приматы», что значит «первые». По-видимому, само это название должно было «научно» засвидетельствовать принадлежность людей к самым совершенным животным. Особенно близки к нам человекообразные обезьяны — антропоиды. Так, человек и шимпанзе имеют общими не менее 99 % своих генов. У нас нет ни одного органа, ни одной структуры, которых бы не было у антропоидов (вплоть до аппендикса). Такое разительное сходство вынуждает нас поставить этих обезьян выше остальных и назвать их высшими приматами. Попробуем взглянуть на высших приматов со стороны, на время забыв о своем явном родстве с ними. Мы обнаружим, что на Земле в настоящее время живет всего несколько видов этих обезьян, ведущих весьма скрытный образ жизни в тропических лесах Африки и Азии. Численность их популяций довольно низка, а для таких видов, как орангутан и горная горилла, даже близка к опасной черте, граничащей с вымиранием. Несмотря на то что в образе жизни и поведении человекообразных обезьян и людей можно отметить множество сходных черт, все же главным предметом нашей гордости — абстрактным мышлением — они, по-видимому, не обладают, а если и обладают, то им явно не пользуются. Поэтому, изумляясь их сходству с нами, мы не можем доказать, почему, например, горилла должна считаться совершеннее леопарда, буйвола или, наконец, любого жука. К тому же, говоря о величии наших животных предков, не надо забывать о других представителях нашего рода Homo. А где они? По свидетельству палеонтологов, данный род никогда не блистал обилием видов, но все-таки когда-то существовали H.habilis (человек умелый) и многочисленные представители H.erectus (человека прямоходящего). Еще совсем недавно (около 50 тысяч лет назад) на Земле обитал близкий нам вид (или, может быть, даже подвид) — неандерталец (Homo sapiens neanderthalensis). Между прочим, по величине головного мозга он нам не уступал, а особенности захоронений явно указывают на его способность к абстрактному мышлению. И тем не менее неандерталец вымер, по-видимому, не выдержав борьбы за существование. Конечно, можно фантазировать вокруг непосредственных причин этого события; например, можно вообразить, что именно наш вид повинен в гибели неандертальцев, или же, что мы просто поглотили их при метисизации (межрасовой гибридизации). Наконец, вспомним свое еще совсем недавнее прошлое. Что представлял собой Н. sapiens всего каких-нибудь 15 тысяч лет назад? Небольшие группы (по 20–30 человек) кочующих охотников-собирателей, затерянные в огромных пространствах девственных лесов, не производят впечатления отважных покорителей природы. Присмотритесь к аборигенам Австралии или Амазонии, послушайте их мифы и легенды и вы отметите, с каким уважением и прямо-таки благоговением относятся они не только к крупным млекопитающим — основным объектам своей охоты, но даже к совсем мелким зверькам, птицам и пресмыкающимся. Да и наши сказки и поверья изобилуют взаимными превращениями человека, животных и растений друг в друга. Человек в этих историях предстает перед нами совсем не гордым исполином, а скромным просителем у духов удачи в весьма ненадежном деле охоты. Положение круто изменилось с появлением земледелия и скотоводства. Контроль над разведением некоторых видов животных и растений дал человеку возможность ощутить свое превосходство над ними. Новая точка зрения на природу получила отражение в Ветхом Завете, где Бог говорит только что созданным людям: «Владычествуйте над рыбами морскими, и над зверями, и над птицами небесными, и над всяким скотом, и над всею землею, и над всяким животным, пресмыкающимся на земле». Таким образом, нашему бытовому антропоцентризму не более 8—10 тысяч лет. Дальнейший ход истории сопровождался непрекращающимся техническим прогрессом, основу которого составляло совсем не стремление к всеобщему благу, а гораздо более прозаичная так называемая престижная экономика. Представители рода человеческого в большинстве своем стремились и стремятся не столько к удовлетворению своих нужд в пище, крове и т. д., сколько к более высокому положению на социальной лестнице. Главными «объективными» аргументами в людском обществе обычно выступают близость к «аристократическим» родам и так называемое богатство. Наш спортивный, соревновательный дух, вполне оправданный в первобытные времена, теперь мешает избавиться от таких пороков, как вещизм, национализм и… антропоцентризм. Высшие и низшие Любой современный образованный человек, конечно, знает, что окружающий его мир живых организмов является продуктом естественного процесса — эволюции. Он также знает, что теорию эволюции создал Ч. Дарвин. Из нее следует, что каждое последующее поколение приспособлено к жизни немного лучше предыдущего, т. е. оно в некотором смысле более совершенно. В итоге в ходе эволюции должно наблюдаться непрекращающееся совершенствование всех форм жизни.Экклесиаст, III, 21*
 Рис. 1. Историческое развитие царства животных (по: [Мир живой природы, 1984]).
Рис. 1. Историческое развитие царства животных (по: [Мир живой природы, 1984]).
Как мы знаем из школьной программы, где-то в ордовике на нашей планете появились рыбы (см. таблицу и рис. 1), затем в девоне — карбоне от них ответвились амфибии, которые приступили к освоению суши. Земноводные породили пресмыкающихся, а те за мезозойскую эру воплотились в огромное число форм, среди которых всегда вспоминают динозавров. Затем динозавры, а с ними и другие монстры (птеродактили, ихтиозавры и пр.) в конце мелового периода вымерли, уступив планету более прогрессивным птицам и млекопитающим. Последние, быстро совершенствуясь, породили высших приматов. Наконец, около 15 миллионов лет назад появились первые представители семейства Hominidae. Эволюция гоминид протекала под знаком увеличения головного мозга, совершенствования речи и орудий. Быстро промелькнули стадии: рамапитек, австралопитек, человек способный, человек прямоходящий (питекантроп, синантроп, гейдельбергский человек), неандерталец и, наконец, наш вид (или подвид) — кроманьонец. Ура! Мы победили всех, потому что именно мы завершили эту жуткую гонку. Наше совершенство заслуженно и не вызывает сомнений. Геохронологическая шкала
| Эон | Эра | Период | Изотопные датировки, млн лет | Основные события и доминирующие группы |
|---|---|---|---|---|
| Фанерозой | Кайнозой | Четвертичный | 1,8 | Человек |
| Неоген | 25 | Млекопитающие, птицы, насекомые | ||
| Палеоген | 66 | Млекопитающие, птицы, насекомые | ||
| Мезозой | Мел | 136 | Вымирают динозавры и аммоноидеи; Аммоноидеи, динозавры | |
| Юра | 190-195 | Первые птицы; Аммоноидеи, динозавры | ||
| Триас | 230 | Первые млекопитающие; Первые динозавры; Рептилии | ||
| Палеозой | Пермь | 280 | Рептилии | |
| Карбон | 345 | Первые рептилии; Земноводные | ||
| Девон | 400 | Первые земноводные; Первые аммоноидеи; Рыбы | ||
| Силур | 435 | Моллюски, брахиоподы | ||
| Ордовик | 490 | Первые рыбы; Брахиоподы, трилобиты, иглокожие | ||
| Кембрий | 570 | Трилобиты | ||
| Поздний докембрий (Венд)) | 650—690 | Кишечнополостные | ||
Однако осмотримся вокруг себя. На Земле в данный момент обитают не менее двух миллионов видов животных, с биологической точки зрения таких же сущностей, как и мы. Получается, что вместе с нами эволюционное соревнование завершили еще два миллиона видов. У каждого из них был свой вид-предок: выходит, что за каждым видом стоит длинная цепь предков, которая растянулась по меньшей мере на два миллиарда лет. Но если рыбы — низшие, т. е. менее совершенные, то почему же они не вымерли, почему и сейчас в морях и пресных водоемах обитает их не менее 20 тысяч видов? Почему вымерли далеко не все рептилии? Даже после страшных катастроф, обрушившихся на Землю в конце мелового периода, современные рептилии по числу видов не уступают млекопитающим. Если считать, что выжившие пресмыкающиеся совершеннее вымерших, то придется признать, что змеи и ящерицы по уровню организации выше динозавров, ихтио- и плезиозавров. С этим не согласится ни один зоолог. О жалкой видовой численности высших приматов мы уже упоминали. И наконец, главная несуразица! Большая часть видов животных приходится на беспозвоночных, из которых на первом месте стоят насекомые. Некоторые семейства жуков или бабочек содержат больше видов, чем тип хордовых со всеми позвоночными (рис. 2).
 Рис. 2. Соотношение числа видов в различных систематических группах современной фауны (по: [Росс и др., 1985]).
Рис. 2. Соотношение числа видов в различных систематических группах современной фауны (по: [Росс и др., 1985]).
Таким образом, если встать на объективную точку зрения, то получается, что победителями в соревновании за жизнь оказались насекомые, а хрестоматийная последовательность: рыбы — земноводные — рептилии — птицы — млекопитающие — человек — это лишь последовательность появления новых форм. То, что суша осваивалась не рыбами, а амфибиями и рептилиями, довольно тривиально. Новое, существующее рядом со старым, может оказаться не лучшим, а просто другим. Таким образом, мы видим, что прямолинейный подход, при котором совершенство связывается с победой в борьбе за существование, вроде бы приводит к парадоксу. Но позвольте, разве не очевидно, что рыбы по уровню внутренней организации уступают зверям и птицам? Разве не очевидно, что инфузории и амебы проще любого червя, а низшие растения вроде мхов много примитивнее пальм и эвкалиптов? Действительно, не прислушаться ли нам к мнению мудрецов, живших до Ч. Дарвина, когда еще не были произнесены магические слова «естественный отбор» и «борьба за существование». Представление о разной степени совершенства живых организмов, населяющих Землю, покоится прежде всего на нашем антропоцентризме. С одной стороны, мы признаем себя частью животного мира, а с другой — четко фиксируем «пропасть», отделяющую человека от остальных животных. Наше кардинальное отличие заключается не в том, что мы лишены шерсти, ходим на двух конечностях и издаем членораздельные звуки, а в том, что мы способны к абстрактному мышлению. Данная способность позволяет людям планировать свои действия, подчиняя их ясно осознаваемым целям. Правда, это же заставляет нас при рассмотрении любого происходящего в природе процесса задавать вопросы: Для чего? С какой целью? Эти вопросы хорошо помогают понять причины поступков других людей, но правомерны ли они во многих других случаях? Например, вопрос: «Для чего солнце светит?» скорее всего вызовет у читателя улыбку, потому что свечение солнца не имеет никакой цели, а ведь еще совсем недавно на этот вопрос отвечали: «Чтобы нам было светло и тепло». Даже такой общепризнанный мудрец всех времен, как Аристотель (384–322 до н. э.), считал, что в природе все «существует ради чего-то или должно соответствовать тому, что существует ради чего-то». Считая целеполагающую деятельность человека высшей формой поведения живых существ, Аристотель первый разделил их по степени совершенства. Ниже всех он поставил растения с их растительной душой, т. е. способностью к росту и размножению. Несколько выше были помещены «низшие» малоподвижные животные, которые способны чувствовать только при контакте (т. е. обладают осязанием, вкусом). Более высокую ступень заняли подвижные животные, способные воспринимать сигналы из окружающего мира также и на расстоянии с помощью зрения, слуха и обоняния. Их стремления не основаны на рассудке, но они способны к воображению. На высшую же ступень было поставлено единственное животное — человек, который кроме воображения обладает еще и рассудком. Восходящий ряд существ строился Аристотелем в соответствии с усложнением их души. Растительной душой обладают все члены ряда, «высшие» животные наделены всеми свойствами души «низших», а человек имеет самую сложную душу. Вроде бы все логично, хотя сами слова «высший» и «низший» вызывают некоторую неудовлетворенность. Пусть даже душа высших состоит из большего числа компонентов, но «больше» не значит «выше». Быть может, причина отождествления усложнения с повышением кроется в своеобразии нашей общественной жизни. Заметим, что у нас, кто командует, тот и выше. Ведь говорим же мы о представителях власти — «верхи». Любой человек, даже на низшей ступени социальной лестницы, может повелевать домашними животными, значит, он их выше. Предположив, что более высокое положение людей в сравнении с другими подвижными животными связано с повышенной сложностью человеческой души, Аристотель продолжил нисходящий ряд к низшим животным и растениям. Как появилась в природе эта лестница живых форм, Аристотель не объясняет, хотя, учитывая особенности его философии, можно предположить, что в основе ее лежали так называемые конечные причины — целефакторы. Под их действием материя постоянно стремится воплотиться в более сложные и более гармоничные формы. Эта идея о стремлении природы к сложной организации стала неотъемлемым элементом большинства философских учений последующих столетий. Научный антропоцентризм Антропоцентрический принцип классификации живых организмов сохранялся в течение двух тысячелетий. Даже у Ж.-Б. Ламарка (1744–1829), создателя первой эволюционной теории, мы читаем: «Можно признать вполне достоверным фактом и очевидной истиной, что из всех видов животных именно организация человека является самой сложной и самой совершенной, как в целом, так и в отношении тех способностей, которыми она его наделяет», и далее — «можно утверждать, что чем больше организация животного приближается к организации человека, тем она сложнее и тем большего совершенства она достигла, и наоборот: чем дальше данная организация от человека, тем она проще и тем менее совершенна». Эволюционная теория Ламарка предполагает, что заполнение ступеней лестницы живых существ (градация) шло постепенно снизу вверх под действием особой «силы природы». Этой таинственной силе препятствовала некая «изменяющая причина», нарушающая строгое соблюдение принципа градации. Заметим, что Ламарк отождествляет усложнение с совершенствованием, поэтому, по его мнению, живые существа в процессе эволюции изменяются от более простых к более сложным и одновременно от более примитивных к более совершенным. По его представлениям, самые примитивные и простые возникают из неживой природы путем самозарождения. Мы видим, что эволюционное учение Ламарка не очень-то помогает в ответе на вопрос, как мы подразделяем живые организмы по степени совершенства. Сходство с человеком, даже отвлекаясь от искусственности этого критерия, трудно использовать при оценке степени совершенства беспозвоночных — все они в равной мере далеки от человека. Интересно, что, несмотря на всю нелогичность антропоцентризма, он продолжает жить даже среди современных антропологов. Так, Я.Елинек в «Большом иллюстрированном атласе первобытного человека» пишет: «Человек, или по-научному Homo sapiens, самое совершенное из всех живых существ». Как это ни странно, но крупнейший вклад в объективное обоснование лестницы существ сделал ярый противник эволюционной идеи знаменитый Ж.Кювье (1769–1832). Он хотел построить классификацию животных только на фактах, при этом пытался найти признак, наиболее характерный для данного царства живых существ. Таким признаком, по мнению Кювье, является строение нервной системы, которая «… по существу и есть все животное; другие системы существуют лишь для того, чтобы ее поддерживать и обслуживать». Соответственно все животные разбиваются на четыре типа — позвоночные, моллюски, членистые и лучистые. Фактически Кювье материализовал то, что Аристотель считал душой. Выбрав признак, якобы лучше всего передающий сущность животного организма, он, точно следуя великому греку, видит ее в развитии психики и двигательной активности. При всем уважении к Кювье согласиться с делением органов на более и менее важные трудно. Ведь хорошо известно, что все системы организма в равной степени нужны для поддержания его жизни. Повреждения как мозга, так и сердца, почек, кишечника одинаково «надежно» ведут к болезни и гибели. Да и сам Кювье считал, что «каждый организм образует единое замкнутое целое», и «что если какая-либо из функций организма будет изменена вне соответствия с изменением других отправлений организма, то он не сможет существовать». Нельзя не согласиться с тем, что нервная система вносит большой вклад в интеграцию организма, однако в этом ей едва ли уступает, к примеру, кровеносная система, и мы не в состоянии понять, какая из них для животных важнее. А разве вклад почек в целостность организма меньше? Создается впечатление, что нервная система выбрана в качестве «объективного» критерия только потому, что головной мозг достигает максимального развития у человека. Таким образом, выбор в качестве критерия высоты организации сложности души или типа нервной системы в равной мере обусловлен антропоцентризмом. Дарвиновская идея естественного отбора основана на представлении о различиях в приспособленности конкурирующих друг с другом особей. «Переживание более приспособленных» совсем не должно автоматически означать выживание более сложных или тем более прогрессивных. В связи с этим не удивительно, что один из ближайших сподвижников Дарвина Т.Хаксли (1825–1895) относился к самой идее эволюционного прогресса весьма скептично. Особое впечатление произвели на него так называемые «персистентные формы», т. е. виды или роды, практически не изменившиеся за десятки и даже сотни миллионов лет. Эти формы весьма нередки среди моллюсков, ракообразных, иглокожих и брахиопод. Достаточно вспомнить о щитне Triops — листоногом ракообразном, сохранявшим свою родовую (а может быть, и видовую) принадлежность более 200 миллионов лет. Т. Хаксли считал, что все современные организмы одинаково хорошо приспособлены к своей среде, и деление на высших и низших нельзя признать научным. Отношение к этой проблеме самого Ч. Дарвина отражает фрагмент его письма Дж. Гукеру от 30 декабря 1858 г.: «…я недумаю, чтобы у кого-нибудь было ясное представление о том, что подразумевается под „высшим“…». Большинство современных дарвинистов вообще считают деление на «высших» и «низших» анахронизмом и весьма вольно обращаются со словом «прогрессивный». Так, один из творцов синтетической теории эволюции Дж. Симпсон называет таксон более прогрессивным, просто если он возник в эволюции позже. Примерно так же обращается с термином «прогрессивный» и другой крупный американский эволюционист В. Грант. Интересно, что внук Т. Хаксли Дж. Хаксли оказался ревностным приверженцем идеи эволюционного прогресса и реанимировал ее в среде дарвинистов. Дж. Хаксли привлек внимание эволюционистов к явлению смены господствующих (доминантных) форм. Как повествует палеонтологическая летопись, в ордовике в море доминировали моллюски, трилобиты и брахиоподы; в девоне первенство захватывают рыбы; на суше в течение всего мезозоя господствуют рептилии; третичный период — царство птиц и млекопитающих и, наконец, в четвертичном — власть над природой переходит к человеку (см. таблицу). Дж. Хаксли пытался найти объективные критерии эволюционного прогресса. По его мнению, это: 1) повышение скорости метаболизма; 2) усиление заботы о потомстве; 3) увеличение скорости реакции на сигналы внешней среды; 4) возрастание способности управлять средой и, главное, уменьшение зависимости от нее. Четвертый пункт выглядит весьма романтично, но именно ему Дж. Хаксли придает важнейшее значение. Особенно странно звучит тезис об управлении средой, что теоретически возможно лишь для человека. Но о каком управлении может идти речь? Пока мы природу только «покоряем». Еще четче антропоцентризм Дж. Хаксли просматривается в его идее неограниченного прогресса, ведущего к человеку. Вполне традиционно особое внимание уделено развитию центральной нервной системы как главному средству усиления независимости от окружающей среды. Идея неограниченного прогресса была развита советским эволюционистом К.М. Завадским в его представлении о «магистральном прогрессе», ведущем к человеку, т. е. снова подтверждена точка зрения Аристотеля. Еще ярче это сделано П. Тейяром де Шарденом — французским палеонтологом и философом, одним из открывателей синантропа, человеком, искавшим компромисс между наукой и религией. Он писал: «… отбрасывая всякий антропоцентризм и антропоморфизм, я считаю, что существует направление (sens) и линия прогресса жизни столь отчетливо, что их реальность, как я убежден, будет общепринята завтрашней наукой». Он ищет «нить Ариадны», ведущую к выходу из лабиринта разнородных эволюционных тенденций. Как и следовало ожидать, этой нитью для него оказалась степень развития нервной системы и психики. Проделав этот старый трюк, Тейяр де Шарден восклицает: «Как только в качестве меры (или параметра) эволюционного феномена берется выработка нервной системы, не только множество родов и видов строятся в ряд, но вся сеть их мутовок, их пластов, их ветвей вздымается как трепещущий букет. Распределение животных форм по степени развитости мозга не только в точности совпадет с контурами, установленными систематикой, но оно придает древу жизни рельефность, физиономию, порыв, в чем нельзя не видеть признака истинности. Такая стройность, к тому же непринужденная, неизменно постоянная и выразительная не может быть случайной». Конечно, нельзя не признать, что в линии, ведущей от рыб к высшим приматам, действительно шло увеличение относительных размеров головного мозга, особенно его передних отделов. Однако у нас нет никаких оснований придавать особое значение именно этой тенденции. В ряду рыбы — млекопитающие прогрессивному укрупнению и усложнению кроме мозга подверглись сердце с системой сосудов, почки, легкие, структуры поддержания стабильности внутриорганизменной среды. Но в этом же ряду можно отметить тенденцию к снижению репродуктивного потенциала, что, впрочем, компенсируется переходом к живорождению и усилением заботы о потомстве. У нас есть все основания считать прогрессом выработку адаптаций к водному образу жизни предками ластоногих и китообразных. Эти адаптации включают в себя преобразования конечностей, кожных покровов, зубов и т. д. Особенно впечатляет возникновение у китообразных совершенно новой для млекопитающих системы эхолокации при мощном развитии соответствующих разделов головного мозга. То же можно сказать о прогрессивном развитии адаптаций к жизни в воздухе у предков летучих мышей. И, наконец, почему бы не говорить о прогрессивном развитии системы размножения у многих плоских червей, ступивших на путь паразитизма. Обычно при рассмотрении приспособлений к паразитическому образу жизни большее внимание обращают на явление деградации (регресса) многих структур, необходимых организму при свободном образе жизни. Однако трудно серьезно говорить о деградации, когда предковые формы (несмотря на свободный образ жизни) были очень простыми крошечными червячками, которые плавали с помощью ресничек. Следует заметить, что биологическое процветание человека в недавнем прошлом в немалой степени сдерживалось именно этими паразитами. Еще и сейчас во многих районах земного шара «венец всего живущего» поражен ими практически на 100 %. Да и как бороться с таким червем, как, например, цепень? Его тело составлено из многих тысяч вполне автономных члеников. Центральным органом каждого членика является матка, содержащая до 175 тыс. яиц. За сутки цепень, находящийся в человеке, выделяет до 5 млн яиц. Итак, если отбросить антропоцентризм, у нас нет объективных оснований особо выделять прогрессивное развитие центральной нервной системы и мы должны считать его лишь частным случаем широко распространенного явления — укрупнения и усложнения в ходе длительной эволюции какого-нибудь органа или системы органов. Мы постараемся дать рациональное объяснение этой эволюционной тенденции. Ламаркизм Первая теория, объясняющая лестницу существ с эволюционных позиций, была создана великим французским биологом Ж.-Б. Ламарком (1744–1829). Основные ее положения Ламарк сформулировал в четырех законах: «Первый закон. Жизнь своими собственными силами непрерывно стремится увеличить объем всякого наделенного ею тела и расширить размеры его частей до предела, ею самой установленного. Второй закон. Образование нового органа в теле животного является результатом новой появившейся потребности, которая продолжает оставаться ощутимой, а также нового движения, порожденного и поддерживаемого этой потребностью. Третий закон. Развитие органов и сила их действия всегда соответствуют употреблению этих органов. Четвертый закон. Все, что было приобретено, запечатлено или изменено в организации индивидуумов в течение их жизни, сохраняется путем воспроизведения и передается новым индивидуумам, испытавшим эти изменения.» Для Ламарка истинность этих законов самоочевидна. Обратимся к первому закону. Разве мы не наблюдаем рост организма в ходе его развития? Разве это не свидетельствует о существовании особой «ростовой силы»? Во втором и третьем законах канонизируются всем хорошо известные факты повышения эффективности многих органов при их упражнении. На самом деле, если у человека возникла потребность в быстром беге, он может с помощью тренировок существенно улучшить свои спортивные показатели. При этом в его скелете, мышцах, системах дыхания и кровообращения произойдут вполне адекватные, т. е. целесообразные изменения. Почему бы не предположить, что в процессе тренировки человек направляет свою ростовую силу к органам с недостаточной функцией? Получается, что распределение в организме ростовой силы находится под контролем психики. То, что люди (и тем более животные) не вполне осведомлены о деталях собственного строения, не имеет значения, поскольку направлять ростовую силу (в форме потока особых материальных частиц — флюидов) можно совершенно бессознательно. Однако четвертый закон вызывает, мягко говоря, недоумение. Где Ламарк наблюдал его проявление? Неужто источником служила только «народная мудрость», вроде записанной А.С. Пушкиным в его «Сказке о мертвой царевне и о семи богатырях». Там есть место, где мачеха, негодуя на белизну лица падчерицы, восклицает:
«Вишь, какая подросла!
И не диво, что бела:
Мать брюхатая сидела,
Да на снег лишь и глядела!»
Глава 2. Морфофизиологический прогресс
Пока нет не то [что] строгого или точного, но даже мало-мальски приемлемого, разумного, логичного понятия прогрессивной эволюции. Биологи до сих пор не удосужились сформулировать, что же такое прогрессивная эволюция. На вопрос — кто прогрессивнее: чумная бацилла или человек — до сих пор нет убедительного ответа.Морфологический подход Есть два понятия, на которых основана идея прогресса, — сложность строения и высота организации. Мы считаем объект сложным, если в нем удается выделить достаточно стабильные неоднородности, которые можно описывать и экспериментально исследовать. И наоборот, простой объект выглядит однородным и бесструктурным, и изучать в нем в общем-то нечего. Что же люди имеют в виду, когда говорят о высоте организации? В философском словаре можно прочесть, что организация это «внутренняя упорядоченность, согласованность взаимодействия более или менее дифференцированных и автономных частей целого, обусловленная его строением». Данное определение хорошо передает сложность и противоречивость этого важнейшего для нашей темы понятия. Как это строение частей может дать информацию об их взаимодействии? Создается впечатление, что мы a priori должны располагать сведениями о функциональном назначении «более или менее дифференцированных и автономных частей» организма. Представим себе фантастическую ситуацию. Пусть нам дали для исследования экземпляр какого-нибудь инопланетного существа и ничего не сообщили об условиях его обитания. Заметим, что данная ситуация не так уж далека от той, с которой имеет дело современный палеонтолог. Только он обычно располагает лишь отпечатком животного на камне. Развивая фантазию, предположим, что для исследования инопланетных существ можно пользоваться только хирургическим инструментарием и световым микроскопом. Тем самым мы имитируем условия работы биологов вплоть до середины XX века. В результате проведенного исследования будет получена информация о внешней морфологии животного, а также о его анатомии, гистологии и цитологии. Сначала мы изучим форму тела и покровы животного, выделим некоторые обособленные образования и условно назовем их наружными органами, после чего начнем гадать о их назначении. Если форма тела окажется близкой к шару, то мы предположим, что организм обитает в какой-то сплошной среде, скорее всего, в воде. Если из тела будут торчать какие-нибудь выросты, то проинтерпретируем их как органы движения или добычи пищи. Ну а если найдутся два отверстия, то одно из них, конечно, объявим ротовым, а другое — анальным. Вскрыв иносущество, мы, вероятно, найдем в нем более или менее обособленные структуры — внутренние органы. Они могут выглядеть или как сплошные, компактные образования, или как длинные трубки, или как плоские складки и т. д. Разрезав эти органы, мы, почти наверняка, выявим в каждом из них определенным образом расположенные детали. Наконец, с помощью микроскопа мы, конечно, откроем еще более тонкие подробности, для описания которых потребуются новые образы и сравнения. Главная цель нашего исследования будет заключаться в том, чтобы выяснить, как все это работает. Но добьемся ли мы разрешения этой задачи? Представьте на момент Аристотеля, ломающего голову над устройством телевизора! Совершенно очевидно, что главную задачу мы не решим, но можно не сомневаться, что будет создана новая наука, какая-нибудь инобиология, и сотни энтузиастов примутся препарировать, классифицировать и описывать на всех уровнях — от морфологического до молекулярного — устройство этих организмов. И тогда выяснится, что для описания одних требуется много слов (для терминов, сравнений и образов), а для других — мало. Предположим, нам попался иноорганизм, представляющий собой просто шар, заполненный каким-то однородным мелкозернистым материалом. Ясно, что для его описания будет использовано мало слов. Другое дело, если организм имеет вид какого-нибудь усеченного конуса с торчащими во все стороны суставчатыми придатками разной величины и формы. А после его вскрытия обнаружится множество внутренних органов, также состоящих из разнообразных частей. Понятно, что для описания такого существа потребуется много слов. Инобиологи наверняка бы сказали, что второй организм гораздо сложнее первого. Не приходится сомневаться, что в результате такого анализа возникла бы теория, по которой сложные организмы имеют более высокую организацию. И все это при полном отсутствии какой-либо информации о функциональном назначении изученных структур! Имела бы эта теория научную ценность? Трудно сказать. Ведь представление о высоте организации в данном случае должно было возникнуть на основании выявляемых с помощью зрения (пусть даже усиленного электронным микроскопом) неоднородностей. А можем мы различать лишь геометрические структуры, объединяемые в образования более высокого порядка. Если нам удастся выявить такую иерархию структур, то мы найдем и принцип оценки высоты организации. Чем больше ступеней окажется в такой иерархической лестнице, тем выше будет уровень организации. Иерархический принцип устройства структур, по-видимому, имеет универсальное распространение (если, конечно, он не является имманентным свойством нашего сознания). Земля, Солнечная система, Галактика, Метагалактика — все это тоже ступени иерархической лестницы. Средняя плотность вещества во Вселенной столь мала, что существование макротел возможно лишь за счет сил притяжения между их элементами. За подавляющее большинство свойств биологических объектов ответственно электромагнитное взаимодействие. Здесь особо следует выделить один тип такого взаимодействия между органическими молекулами, прежде всего между информационными макромолекулами. Он получил наименование молекулярного узнавания и лежит в основе всех фундаментальных биологических процессов, таких как ферментный катализ, регуляция генной активности, межклеточные взаимодействия и т. д. Уникальное распределение электронных плотностей вдоль молекулы белка способно обеспечить узнавание ею фактически любой другой молекулы (в том числе и такой же, как она сама). Все это открывает путь к последовательному укрупнению молекулярных комплексов. Энергия, освобождающаяся при межмолекулярном взаимодействии, может быть использована для образования или разрыва химических связей, т. е. для осуществления химических реакций. Продукты таких реакций могут быть «узнаны» другими молекулами. Усложнение гомологичных структур Сравнивая организмы, принадлежащие систематически близким видам, легко обнаружить большое сходство во взаимном расположении и строении многих органов. Такие органы, несколько преобразованные в ходе эволюции, называются гомологичными. Например, все млекопитающие обладают головным мозгом, четырьмя конечностями, сердцем, легкими и т. д. Это сходство объясняется единством происхождения сравниваемых видов. Органы, унаследованные от вида-предка, со временем изменялись, хотя, как правило, эти изменения касались лишь величины и формы, не нарушая основного плана взаимного расположения гомологичных частей. Гомологичные структуры, в которых можно выделить повторяющиеся элементы, допускают ранжировку по числу таких элементов. Ярким примером здесь могут служить ряды одномерных структур — линий, различающихся числом изгибов. Таковы знаменитые лопастные линии аммоноидей, вымершей группы головоногих моллюсков, которыми кишели моря и океаны в течение примерно 300 миллионов лет. В этих линиях обычно удается выявить целую иерархию изгибов (рис. 3). На изгибы первого порядка (как обертоны на волны) накладываются изгибы второго порядка, те же, в свою очередь, могут быть деформированы изгибами третьего порядка и т. д. Следовательно, каждому изгибу первого порядка соответствует серия изгибов более высоких порядков. Такая система позволяет ввести формализацию. Формулы лопастных линий основаны на том, что каждому изгибу самого высокого порядка приписывается его место в иерархии изгибов. Буквенные символы в формуле обозначают первичные изгибы, а цифровые индексы — уровень в иерархии. Чем большим числом изгибов обладает лопастная линия, тем, по мнению палеонтологов, выше ее сложность, поскольку тем больше символов приходится вводить в ее формулу.Н. В. Тимофеев-Ресовский
 Рис. 3. Усложнение одномерных структур на примере лопастной линии аммоноидей разных геологических периодов.
а — в — девон; г — карбон; д — пермь; е — юра; ж — мел.
Рис. 3. Усложнение одномерных структур на примере лопастной линии аммоноидей разных геологических периодов.
а — в — девон; г — карбон; д — пермь; е — юра; ж — мел.
Прекрасным примером усложнения двухмерной структуры могут служить легкие позвоночных (рис. 4, Б). У хвостатых амфибий они представляют собой пару гладкостенных мешков, хотя у ряда видов данной группы земноводных поверхность этих мешков может быть увеличена за счет крупноячеистой сети перекладин. У бесхвостых амфибий такие ячеи подразделяются системой вторичных перекладин на более мелкие ячейки — легочные пузыри. У многих рептилий появляются куда более мощные ячеистые перегородки, выступающие в глубь легочных мешков. От этих перегородок могут отходить перегородки второго порядка. В итоге, легкое приобретает губчатое строение, занимая весь объем бывшего мешка. Очевидно, что усложнение легких связано с увеличением их газообменной поверхности. Параллельно идет усложнение системы трубок — трахей и бронхов, проводящих воздух. Тот же тип геометрического усложнения можно обнаружить и у самых примитивных животных — губок. Здесь традиционно выделяют три ступени прогрессивного развития — аскон, сикон и лейкон (рис. 4, А). Усложнение сопряжено с увеличением доли клеток, добывающих пищу, — хоаноцитов.
 Рис. 4. Усложнение двухмерных структур (по: [Хадорн, Венер, 1989]).
А — усложнение губок: а — аксон, б — сикон, в — лейкон; Б — усложнение легких у позвоночных: а — хвостатые амфибии, б — рептилии, в — млекопитающие.
Рис. 4. Усложнение двухмерных структур (по: [Хадорн, Венер, 1989]).
А — усложнение губок: а — аксон, б — сикон, в — лейкон; Б — усложнение легких у позвоночных: а — хвостатые амфибии, б — рептилии, в — млекопитающие.
Можно выделить следующие типы геометрического усложнения. Для одномерных структур — линий, стержней, тонких трубок и балок — это иерархия изгибов и ветвлений, для двухмерных — иерархия складок, выпячиваний, карманов. В обоих случаях повышается размерность. Одномерная лопастная линия своими изгибами может плотно покрыть двухмерную поверхность стенки раковины. Система одномерных клеточных балок создает трехмерную структуру печеночной паренхимы. То же можно сказать о системе тонких трубочек, укладываемых в спирали. Выпячивания двухмерных структур создают трехмерные структуры губчатого или слоистого строения. Для трехмерных структур усложнение часто сопряжено просто с повторением одних и тех же элементов (нефронов, семенных канальцев, мышечных волокон и т. д.). В итоге растет масса органа. Главный показатель, сопровождающий усложнение структуры, заключается в увеличении интенсивности ее функции. Если такое усложнение наблюдается в ходе исторического развития, то его принято называть «прогрессивным». Усложнение в ходе онтогенеза К проблеме структурной сложности можно подойти и с другой стороны. Обратим внимание на то, что любой многоклеточный организм развивается из одной, хотя и очень большой, клетки — оплодотворенного яйца. Эта клетка, если ее изучать под микроскопом, производит впечатление весьма однородного и бесструктурного объекта. И тем не менее гомогенная масса куриного желтка через 21 день насиживания превращается в цыпленка со всеми атрибутами сложного организма. Суть этого процесса, по мысли К. Бэра, сводится к следующему: «Во время развития из гомогенного и общего постепенно возникает гетерогенное и частное». Еще натурфилософы говорили о параллелизме между индивидуальным развитием высших организмов и лестницей существ. Однако тот же Бэр показал, что на самом деле имеет место не повторение стадий, подобных ступенькам лестницы существ, а сходство начальных стадий развития у организмов одного типа (в смысле Ж. Кювье). У всех систематически близких видов можно обнаружить несколько стадий, когда зачатки гомологичных органов располагаются по отношению друг к другу весьма сходным образом, как бы по одному плану. Так, у всех зародышей позвоночных имеется внутренний скелет в виде продольного тяжа — хорды. Выше нее (дорсальнее) располагается нервная трубка, а ниже (вентральнее) — первичная кишка. Образование первичных зародышевых структур у большинства хордовых протекает удивительно сходно. Возьмем, к примеру, нервную трубку — зачаток центральной нервной системы. У всех хордовых можно видеть, как в однородной эктодерме ранней гаструлы в области ее контакта с первичной кишкой постепенно обособляется узкая продольная полоска клеток — нервная пластинка. Затем она прогибается и, пройдя фазу желобка, замыкается в нервную трубку, отделившись от сросшейся над нею эктодермы. Еще в 1828 г. Бэр пришел к заключению, что, во-первых, едва ли правомочно сопоставлять уровень организации животных, принадлежащих разным типам, и, во-вторых, у представителей одного типа этот уровень можно оценить по числу гистологических и морфологических дифференцировок, т. е. по числу деталей, выявляемых на глаз и под микроскопом при разном увеличении. В основе морфологической дифференцировки лежит несколько фундаментальных процессов, протекающих на клеточном уровне: клеточное размножение, клеточное движение, межклеточное взаимодействие, клеточная дифференцировка и, наконец, клеточная гибель. Пожалуй, самым важным для нашей темы является процесс клеточной дифференцировки. У многоклеточных организмов каждая клетка находится в одном из целого ряда весьма устойчивых дискретных состояний — клеточных типов, или дифференцировок. Клетки разных типов отличаются по огромному числу свойств: скорости размножения, спектру синтезируемых молекул, сродству к другим клеткам или внеклеточным структурам, способу клеточного движения и т. д. Одни из них стремятся агрегировать друг с другом, чтобы образовать шаровидные скопления или тонкие, в один клеточный слой, пласты. Другие, наоборот, как бы избегают контактов, третьи тяготеют к бесклеточным мембранам, а четвертые пытаются соединиться с клетками других типов. Самым важным здесь является момент изменения клеткой своего типа. Это событие может произойти по разным причинам. Клетка может дифференцироваться совершенно самостоятельно (возможно, израсходовав какой-то ресурс или наработав какой-то продукт). Данный путь обычен для самых начальных стадий индивидуального развития, в особенности у животных, которых принято помещать на нижние ветви филогенетического древа. Однако чаще всего клетка дифференцируется под влиянием внешнего сигнала, поступившего от других клеток того же зародыша. Этот тип дифференцировки, весьма характерный для позвоночных, получил название эмбриональной индукции. В данном случае сигнал к изменению дифференцировки передается от клеток индуктора к близко расположенным клеткам ткани, отвечающей на индукцию. Клетки нового типа могут затем сами выступать в качестве индуктора, побуждая к дифференцировке другие клетки, иными словами, возникает целый каскад (иерархия) эмбриональных индукций. Необходимость довольно тесных контактов между обоими участниками индукционного процесса объясняет эволюционный консерватизм взаимного расположения многих эмбриональных зачатков, т. е. сохранение в эволюции основного плана строения зародышевых структур. В связи с этим в качестве еще одной меры сложности организма можно выбрать число клеточных типов, встречающихся на всех стадиях его развития. К сожалению, тщательный подсчет данного числа пока не проведен, хотя ясно, что многообразие клеточных типов у насекомых и позвоночных измеряется сотнями, а у червей и других низших беспозвоночных — только десятками. Самая примитивная группа многоклеточных животных — губки — располагает едва ли одним десятком разных дифференцировок. Таким образом, наше интуитивное представление о повышении структурной сложности организмов в ходе исторического развития жизни на Земле, по-видимому, нас не обманывает. Сериальная гомология В строении многих животных, в том числе и у доминирующих в наше время позвоночных и членистоногих, можно обнаружить весьма сходные морфологические структуры, повторяющиеся вдоль продольной оси организма. Такие структуры называют метамерами, а само явление — сериальной гомологией. У позвоночных метамерами являются позвонки, ребра, сомиты и т. д. У кольчатых червей и членистоногих — это, прежде всего, сегменты. У некоторых кольчатых червей (считающихся примитивными) все сегменты с их наружными придатками поразительно сходны. Лишь два из них — головной и конечный — составляют исключение. У других кольчецов (эволюционно «более продвинутых») можно выделить отделы с несколько иным строением сегментов — так называемые тагмы. У членистоногих дифференциация сегментов (тагмозис) зашла гораздо дальше, особенно у высших ракообразных и насекомых. Сравнивая представителей различных групп кольчатых червей и членистоногих, можно получить плавный ряд возрастающей дифференциации метамеров. При этом можно заметить, как простые наружные придатки сегментов кольчатых червей преобразуются у членистоногих в конечности, состоящие из набора трубчатых члеников. Строение конечностей в разных тагмах может поразительно различаться, отражая их функциональную дифференциацию. Они могут быть ногами, органами чувств, хватательными и ротовыми придатками, органами дыхания, приспособлениями для спаривания и для защиты потомства (рис. 5). Американский палеонтолог Дж. Цисне предложил использовать в качестве меры тагмозиса конечностей количество информации (А), определяемое с помощью формулы Бриллюэна:
 где N — полное число конечностей; Ni — число конечностей с i-й функцией, k — число функций.
Оказалось, что у примитивных ракообразных и трилобитов величина тагмозиса равна 0,5 бит, а у высших ракообразных — 2,5 бит (см. рис. 5). Выходит, что степень морфологической сложности и в данном случае допускает прямое измерение. Мы снова убеждаемся в том, что наше деление на высших и низших имеет некоторое объективное основание.
Рассматривая ряды организмов, составленные в порядке возрастания сложности определенных гомологичных структур, трудно удержаться от вопроса, почему же до сих пор существуют (и зачастую явно процветают) виды с примитивными, мало дифференцированными органами? Например, почему бы улиткам не обладать глазами осьминога? И вообще, почему бы всем органам не быть одинаково хорошо развитыми? Ведь было бы совсем неплохо, если бы люди в придачу к их уму обладали бы еще зрением совы, нюхом собаки, слухом кошки, а заодно и способностью слышать ультразвук, видеть тепло и рожать без мук. Однако еще Аристотель подметил: «…природа дает одной части организма то, что она забирает у другой». Знаменитый Жоффруа Сент-Илер назвал эту закономерность «принципомуравновешивания» органов.
где N — полное число конечностей; Ni — число конечностей с i-й функцией, k — число функций.
Оказалось, что у примитивных ракообразных и трилобитов величина тагмозиса равна 0,5 бит, а у высших ракообразных — 2,5 бит (см. рис. 5). Выходит, что степень морфологической сложности и в данном случае допускает прямое измерение. Мы снова убеждаемся в том, что наше деление на высших и низших имеет некоторое объективное основание.
Рассматривая ряды организмов, составленные в порядке возрастания сложности определенных гомологичных структур, трудно удержаться от вопроса, почему же до сих пор существуют (и зачастую явно процветают) виды с примитивными, мало дифференцированными органами? Например, почему бы улиткам не обладать глазами осьминога? И вообще, почему бы всем органам не быть одинаково хорошо развитыми? Ведь было бы совсем неплохо, если бы люди в придачу к их уму обладали бы еще зрением совы, нюхом собаки, слухом кошки, а заодно и способностью слышать ультразвук, видеть тепло и рожать без мук. Однако еще Аристотель подметил: «…природа дает одной части организма то, что она забирает у другой». Знаменитый Жоффруа Сент-Илер назвал эту закономерность «принципомуравновешивания» органов.
 Рис. 5. Различия в уровне тагмозиса (А) конечностей водных членистоногих.
а — трилобит (h = 0,5); б — речной рак (h = 2,5).
Рис. 5. Различия в уровне тагмозиса (А) конечностей водных членистоногих.
а — трилобит (h = 0,5); б — речной рак (h = 2,5).
Если мы скажем, что высокая степень развития всех структур организму не нужна, то мы просто объясним непонятное неизвестным. Почему за высокое развитие одних органов надо платить недоразвитием других, представляет собой серьезную проблему, едва ли сводимую к одной лишь экономии энергии и вещества. Функциональный подход В этом разделе мы попытаемся ответить на вопрос: для чего существуют органы. Заметим, что в физике вопросы такого рода не задаются. Иное дело техника. Когда мы строим дом, машину, дорогу и т. д., мы всегда руководствуемся определенной целью, т. е. мы прекрасно знаем, для чего работаем. В нашей трудовой деятельности все рационально, мы хотим достигнуть желанной цели с минимальными затратами. Ведь смешно же стрелять из пушек по воробьям. Сам процесс труда обычно удовольствия не доставляет, для человека гораздо важнее цель. Вот что сказал по этому поводу Ф.М. Достоевский: «Мне пришло раз на мысль, что если б захотели вполне раздавить, уничтожить человека, наказать его самым ужасным наказанием, так что самый страшный убийца содрогнулся бы от этого наказания и пугался его заранее, то стоило бы только придать работе характер совершенной, полнейшей бесполезности и бессмыслицы». Для уменьшения энергетических затрат человек создает орудия: сначала дубину и каменный топор, а потом и машины. Сотни тысяч лет орудийной деятельности, естественно, наложили отпечаток на наши мышление и язык. Глаза нам нужны, чтобы видеть, уши — чтобы слышать, руки — чтобы работать, т. е. все органы являются лишь средством для удовлетворения наших желаний. Домашние животные человеку также даны для удовлетворения потребности в пище, для переноса тяжестей и т. д. Более того, той же цели могла бы служить и вся природа. Но, увы, приспособить природу удается далеко не всегда, поэтому для завершения картины мира, где все подчинено целям, надо было придумать богов — носителей целей более высокого порядка, для достижения которых и люди, и все объекты природы выступают как орудия. Поняв это, мы должны были бы отказаться не только отвечать на вопросы «для чего» и «ради чего», но даже и ставить эти вопросы. Более того, следовало бы признать ненаучным и сам термин «функция». Однако немного подумав, мы поймем, что это странное целевое мышление помогает человеку эффективнее организовать свою деятельность для удовлетворения своих насущных потребностей, в конечном счете такое мышление способствует увеличению числа потомков. Но разве не те же потребности испытывают особи любого вида? Тут мы подходим к очень важному пункту нашего рассуждения — сущности живого. По-видимому, главное свойство жизни — это ее принадлежность к открытым термодинамическим системам, находящимся вдали от равновесия. Длительное существование таких систем возможно только за счет постоянного притока извне энергии и вещества. Внутри живой системы идут постоянные процессы старения (повышения ее энтропии), но от гибели ее удерживают процессы омоложения (производства отрицательной энтропии). Иначе говоря, для длительного существования популяции ее особи вынуждены все время питаться и стареть, компенсируя последнее постоянным производством потомства. Кроме того, учитывая смертность не только по причине старения, но и по другим (изменение физикохимических условий среды, хищничество, паразитизм и т. д.), производство потомства должно компенсировать все варианты гибели особей. Теперь приглядимся к тому, что человек делает, когда работает. Возьмем, к примеру, тяжелый труд земледельца. Человек очищает поле от леса, взрыхляет почву, помещает в нее семена злаков, борется с сорняками и пр. Наконец, жнет, молотит, перетирает зерно и печет хлеб. Сколько труда он затратил! Крестьянину кажется, что он выращивал хлеб, а на самом деле хлеб вырастал сам. Да и не в силах хлебопашец осуществить химический синтез углеводов из углекислоты и воды. Это делается самим растением, использующим в качестве источника энергии солнечные лучи. Человек же только создает условия для роста нужного злака. Работая в течение примерно ста дней в году «в поте лица своего», он при самой примитивной технике в состоянии обработать около трех гектаров пашни и получить с них около двух тонн зерна, что более чем достаточно для жизни его семьи в течение года. Таким образом, работа земледельца фактически сводится к отведению части лучистой энергии Солнца к биомассе людей. Однако сам крестьянин ставит перед собой куда более прозаичную цель. Он трудится, чтобы обеспечить себя и свою семью пищей еще на один год. Отметим, что для выживания ему нужно трудиться и обладать знаниями по выращиванию хлеба. Теперь перейдем к рассмотрению абстрактной живой системы. С биологической точки зрения, главное заключается в преобразовании поглощенной энергии и вещества в биомассу потомства, ибо только это дает популяции возможность существовать неопределенно долго. В чем заключается материальная основа этого чуда? Что заставляет вещество из окружающей среды постоянно превращаться в биомассу популяции? Уж не жизненная ли сила? Рассмотрим какое-нибудь элементарное событие в этом потоке энергии и вещества. Ну, скажем, этап распада поглощенных углеводов. Суммарное уравнение, описывающее окисление глюкозы, выглядит очень просто: глюкоза + 6O2 → 6СО2 + 6Н2О + 680 ккал. Эта реакция может идти и сама по себе без всякого участия живых организмов. Например, нагрев сухую глюкозу, мы можем ее сжечь, т. е. точно так же превратить в CO2 и Н2O, но в этом случае вся разность свободных энергий углевода и продуктов его сгорания практически моментально рассеется в форме тепла, не совершив никакой полезной работы. У живых организмов эта энергия освобождается поэтапно, аккумулируясь в специальных химических соединениях (чаще всего это АТФ). Каждая грамм-молекула АТФ запасает около 7,3 ккал, которые затем могут быть использованы для совершения различных видов работы. Поэтому биологический смысл приведенной выше реакции состоит в преобразовании химической энергии глюкозы в энергию нескольких молекул АТФ: глюкоза + 6O2 + 36 АДФ → 6СО2 + 36 АТФ + 42Н2O. Сравнив эту реакцию с предыдущей, мы видим, что живой системе удается использовать только 263 ккал (7,3 ккал х 36) из 680 возможных, т. е. эффективность использования энергии составляет 39 %, остальные 61 % безвозвратно теряются в виде тепла. Нематериальная жизненная сила могла бы иметь к.п.д. и повыше. Рассматривая любой биохимический процесс распада, мы всегда встречаемся с одной и той же ситуацией: запас свободной энергии у реагирующих компонентов выше, чем у конечных продуктов. Это является индикатором необратимости процесса, самопроизвольности его протекания, хотя ничего не сообщает о его скорости. Но именно скорость имеет в данном случае принципиальное значение. Выходит, мудрое изречение «под лежачий камень вода не течет» отражает глубокую истину. Ведь, собственно говоря, как это вода не течет под камень? С научной точки зрения, вполне течет, только очень медленно. Чтобы потекла быстро, камень надо слегка пошевелить, т. е., приложив усилия, совершить работу. Мешок с глюкозой можно хранить годами и, несмотря на свободный доступ кислорода, процесс окисления сахара идет крайне медленно. Оказывается, для запуска любой химической реакции реагенты должны преодолеть некоторый энергетический барьер (иной раз довольно существенный). Химик решает эту проблему, нагревая реакционную смесь или внося в нее катализатор. Живые организмы идут по второму пути. Биохимические реакции ускоряются с помощью специальных катализаторов — ферментов — белков, способных образовывать специфичные комплексы с молекулами реагентов. Ферменты не вносят в систему энергию, а только снижают активационные барьеры реакций. Как это происходит — предмет специальной науки — молекулярной энзимологии. Однако ясно одно, ничего сверхтаинственного здесь нет. Энергия, необходимая для изменения пространственной конфигурации реагирующих молекул, освобождается при образовании специфичного комплекса с ферментом. Каталитические свойства фермента всецело обусловлены строением его молекулы, которое, в свою очередь, полностью определяется последовательностью нуклеотидов ДНК соответствующего гена. Следовательно, в данном случае продукт гена способен ускорять протекание вполне определенной химической реакции. Выражаясь более антропоморфно, можно сказать, что организм, располагая таким геном, обладает «знанием» о том, как осуществить совершенно конкретную химическую реакцию. Это знание, очевидно, заключено в последовательности нуклеотидов генного локуса. Чтобы знание подобного рода «сработало», нуклеотидная последовательность гена должна быть преобразована в аминокислотную последовательность белка, размноженную в гигантском числе экземпляров, т. е. на синтез молекул фермента нужно затратить немало энергии и вещества. Итак, живая система, с одной стороны, должна обладать информацией о строении белка, а с другой — затратить энергию на его синтез и при этом, очевидно, остаться в выигрыше. Синтез макромолекул сам собой не идет. Для его осуществления мало располагать ферментами, нужна еще и энергия. Самопроизвольное протекание реакции синтеза можно обеспечить ее сопряжением с реакцией распада, наподобие рассмотренной выше. Нужно, чтобы свободная энергия исходных реагентов обеих реакций — сопрягающей и сопрягаемой — была выше свободной энергии их продуктов. Опыт биохимии свидетельствует, что этот энергетический закон выполняется во всех без исключения случаях. Получается, что жизнь как явление вполне подчиняется законам химической термодинамики. Это отражено в одном из определений жизни, данном Ж. Перре: «Жизнь есть потенциально способная к самовоспроизведению открытая система сопряженных химических реакций, катализируемых последовательно и почти изотермично сложными и специфичными органическими катализаторами, которые сами вырабатываются этой системой». Итак, все макромолекулы организма (в том числе и все ферменты) синтезируются за счет энергии, поступающей из внешней среды. Для животных источником этой энергии обычно служат белки, жиры и углеводы других организмов. Для растений таким источником является Солнце. В этом отношении все живые системы равны, их различия касаются только их динамических характеристик и прежде всего скорости синтеза биомассы. Функции Прямой анализ нуклеотидных последовательностей ДНК выявил принципы внутреннего устройства генов. Оказалось, что огромное большинство генов кодирует аминокислотные последовательности белков, обладающих в принципе безграничными возможностями молекулярного узнавания. В одном случае это ферменты, узнающие молекулы субстрата и снижающие активационные барьеры соответствующих химических реакций; в другом — так называемые структурные белки, занимающие свои совершенно определенные места в каких-то мультимолекулярных ансамблях; в третьем — белки-регуляторы, узнающие определенные последовательности ДНК. Во всех случаях нуклеотидная последовательность, кодирующая первичную структуру белка, фактически определяет его функцию. Будем считать, что структура обладает функцией, если она обеспечивает протекание любого процесса, способствующего преобразованию энергии окружающей среды в биомассу популяции. Такие структуры будем называть рабочими. Оказалось, что все молекулярные структуры, закодированные в генах, обладают какой-то функцией. Более того, большинство этих функций настолько важны, что удаление гена очень часто вызывает остановку развития организма. Теперь попробуем разобраться с предназначением видимых глазом морфологических структур. Возьмем, к примеру, такую структуру, как зубы позвоночного. Их очевидная функция состоит в том, чтобы питательное вещество, заключенное в относительно крупной жертве, быстрее обработать ферментами пищеварительного тракта. Тонкое измельчение пищевого куска повышает на порядки скорость его переваривания. Ну, а если размер пищевого куска больше самого хищника, то без зубов и челюстей вообще не обойтись. Получается, что челюсти с зубами выполняют функцию, сходную с пищеварительными ферментами: и те, и другие ускоряют процесс первичной обработки пищи. Челюсти действуют здесь как катализатор дезинтеграции тела жертвы. Для этого требуется энергия, которая идет на механическую работу измельчения, а также на создание и поддержание соответствующих рабочих структур. Вообще, для поддержания жизни должен быть выполнен целый ряд объективно необходимых условий. Процесс питания включает в себя несколько стадий, неумолимых в своей последовательности. Особь должна встретиться с источником пищи (обычно это особь другого вида), измельчить ее, химически расщепить макромолекулы жертвы до униформных блоков и, наконец, доставить данные блоки каждой своей клетке. Заметим, что вся эта сложная цепь оборвется, если нарушится хотя бы одно ее звено. Само собой разумеется, что для существования организма должно постоянно выполняться условие его охраны от негативных внешних факторов, зачастую весьма агрессивных. Это достигается с помощью панцирей, шипов, средств оптической маскировки, иммунной защиты и т. д. Среда, в которой существует живая система, изменчива, флуктуируют все ее параметры: температура, влажность, соленость и т. д. Все это могло бы нарушить течение многих жизненно важных процессов. Поэтому организм должен обладать способностью сохранять значения своих внутренних параметров при флуктуациях среды, иными словами, он должен обладать системой внутреннего гомеостаза. Кроме того, особь может погибнуть, что называется, во цвете лет просто случайно (от удара молнии, пожара, потопа, кирпича, упавшего с крыши, и т. д.). Есть только одно средство скомпенсировать ущерб от всех этих напастей — особи популяции должны постоянно воспроизводиться. Но это тоже не очень-то просто: ведь необходимо произвести половые клетки (гаметы), обеспечить их слияние и защитить от внешней среды первые стадии онтогенеза. Итак, для неопределенно долгого (стационарного) существования живой системы необходимо, чтобы: 1) извне в нее постоянно поступали энергия и вещество, 2) из нее во внешнюю среду постоянно удалялись тепло и отработанные продукты, 3) особи охранялись от преждевременной гибели, 4) внутренние параметры системы были стабильными, 5) шло постоянное производство новых особей. Перечисленными свойствами должны обладать любые многоклеточные организмы, иначе говоря, все они должны обладать пятью основными функциями: доставки, удаления, охраны, гомеостаза и воспроизведения. Если исследовать все многообразие взаимодействий между организмом и средой, а также между различными частями одного организма, то нам откроется бесчетное число функций, подчиненных основным. Возьмем, к примеру, кости конечностей позвоночных. Мы найдем здесь и поверхности суставов, и отростки для прикрепления мышц, и отверстия для нервов и сосудов, т. е. морфологические признаки, имеющие четкую функциональную нагрузку. Распилив кость, обнаружим, что костные балки располагаются вдоль линий сжатий и растяжений, в полном соответствии с теорией сопротивления материалов. Кроме того, в полостях трубчатых костей млекопитающих мы найдем кроветворную ткань — костный мозг. А у птиц эти полости заполнены воздухом и связаны с легочными мешками; получается двойной функциональный эффект — масса тела уменьшается, а дыхание интенсифицируется. Подобный анализ можно продолжать бесконечно, и всюду мы будем находить функциональное назначение морфологических структур. Этот поиск и обнаружение целесообразности в строении живых организмов настолько завораживают, что возникает невольная мысль о целесообразности их малейших черточек. Такая точка зрения, безусловно, относится к ненаучным, так как в реальном мире нет места для идеальных объектов. Совершенно ясно, что доведенная до предела целесообразность автоматически требует признания существования бога. Обобщив результаты нашего функционального анализа, можно прийти к заключению, что живое вещество фактически является лишь средством для жизни как процесса, состоящего в производстве этого же вещества, т. е. цель и средство здесь удивительным образом переплетены. Получается, что для потребления энергии из окружающей среды живая система должна обладать большим набором функций и постоянно затрачивать энергию на синтез соответствующих рабочих структур. Вообще, живое вещество можно рассматривать как единый катализатор, способный сам себя синтезировать в процессе катализируемой реакции. Ясно, что масса живого вещества будет расти в геометрической прогрессии, и в той же прогрессии будет нарастать энергетический поток внутрь живой системы. Предел этому может положить только ограниченность ресурсов внешней среды. Итак, любая живая система (будь то людское общество или природная популяция) для продления своего существования должна постоянно потреблять из окружающей среды энергию и вещество. Большая часть этого потока растрачивается на тепло и отбросы, а остальная идет на создание и поддержание рабочих структур, сохраняющих мощность энергетического потока вовнутрь системы на прежнем уровне. Каждая такая структура выполняет свой, специфичный для нее вид работы. Мерой функциональной эффективности структуры может служить ее мощность, т. е. объем работы, выполняемой за единицу времени в режиме максимальной нагрузки. Нередко структура является необходимой, но инертной частью какого-нибудь сложного аппарата (например, локомоторного), или же ее функциональное назначение связано с пассивной защитой (например, панцирь с шипами). В этом случае строение структуры должно соответствовать максимальной нагрузке со стороны каких-то активных внешних или внутренних факторов. Способность уравновешивать подобные нагрузки также можно назвать мощностью. Это позволяет говорить о мощном костяке, мощных крыльях, мощном панцире и т. д. Достаточно беглого взгляда на рис. 4, Б, чтобы сказать, что по мощности легких млекопитающие значительно превосходят земноводных. Выходит, что из анализа строения ряда гомологичных структур мы можем извлечь информацию об их функциональном назначении и мощности. В целом же все рабочие структуры — от молекулярных до морфологических — можно считать информационными. Выражаясь более антропоморфно, любая живая система обладает «знанием» об условиях преобразования энергии и вещества внешнего мира в собственную биомассу. Аллометрия Родственные виды животных обычно различаются средними размерами своих особей. Кроме того, типичные особи могут различаться по многим количественным характеристикам (параметрам), таким как величина любого органа, потребление кислорода, число митохондрий в клетке печени, средняя продолжительность жизни и многое другое. Оказалось, что для совокупности видов одного таксона величины подобных параметров (X) очень часто связаны с массой особи (М) так называемой аллометрической зависимостью X = а*Мα, (2.2) где а и α — аллометрические коэффициенты. Заметим, что зависимости такого рода связывают две характеристики типичной особи каждого вида в пределах всей таксономической группы. Известно, что при увеличении размеров шара его объем возрастает пропорционально кубу, а поверхность — только квадрату радиуса. Этот же закон обычно выполняется и при пропорциональном (изометричном) увеличении тел более сложной формы: длины всех одномерных структур возрастают пропорционально кубическому корню из массы тела, т. е. пропорционально М0,33, а площади всех поверхностей — пропорционально М0,67. Выберем в качестве параметра одну из важнейших характеристик особи — скорость потребления ею энергии в состоянии покоя, т. е. мощность основного метаболизма (обмена веществ). Этот параметр можно оценить либо по скорости потребления кислорода, либо по скорости рассеивания тепла. Естественно, при увеличении размеров особи растет и мощность ее метаболизма (Р). Эта связь выражается эмпирическим уравнением Р = а*М0,75. (2.3) Здесь коэффициент а численно равен мощности основного метаболизма особи с единичной массой (скажем, с массой 1 кг). После логарифмирования уравнение (2.3) приобретает вид ln Р = ln а + 0,75 ln М. (2.4) Значит, в логарифмическом масштабе мы должны иметь линейную связь между мощностью метаболизма особи и массой ее тела. Если эту связь представить графически, то все плацентарные млекопитающие (от мыши до слона) и птицы (кроме воробьиных) попадут на одну прямую, наклон которой равен 0,75 (рис. 6). Все эти животные имеют очень близкие значения коэффициента а. Для сумчатых, плацентарных млекопитающих и воробьиных птиц данный коэффициент равен соответственно 46,5; 73,3 и 129,0. Для амфибий и рептилий его значения колеблются в диапазоне от 0,8 до 7,8. Получается, что особи одной массы, но принадлежащие к разным таксонам, рассеивают в состоянии покоя разное количество энергии.
 Рис. 6. Кривая «от мыши до слона». Зависимость мощности метаболизма от массы у млекопитающих и птиц (по: [Benedict, 1938]).
Рис. 6. Кривая «от мыши до слона». Зависимость мощности метаболизма от массы у млекопитающих и птиц (по: [Benedict, 1938]).
Разделив обе части уравнения (2.3) на М, получим связь массы тела со скоростью рассеивания энергии единицей массы животного, т. е. с удельной мощностью метаболизма (р): р = а*М-0,25. (2.5) Знак минус при показателе степени свидетельствует о том, что с ростом массы особи удельная мощность метаболизма падает. Эта закономерность верна для всех животных, объединенных общим значением коэффициента а. Заметим, что у млекопитающих размеры гомологичных клеток отличаются незначительно, поэтому уравнение (2.5) указывает на ослабление метаболизма средней клетки при увеличении массы животного. Жизнь особи многообразна. Животное может и мирно отдыхать, и мчаться изо всех сил, спасаясь от смертельной опасности или преследуя свою жертву. Совершенно ясно, что в течение своей жизни организм неоднократно испытывает максимальные физические нагрузки. Главные энергетические расходы животных, безусловно, связаны с активным движением. Достаточно отметить, что при интенсивных мышечных усилиях потребление кислорода возрастает в 10–30 раз. Поэтому мы должны прежде всего рассмотреть, как изменяется режим работы мышц при увеличении размеров особи. Как известно из механики, работа (A) равна произведению силы на путь. При сокращении мышцы на длину Δl она совершает работу в соответствии с формулой А = S*σ*Δl, (2.6) где S — площадь поперечного сечения мышцы; σ — сила, развиваемая мышечным пучком единичного сечения. Обширный экспериментальный материал свидетельствует, что у всех животных величина σ очень близка. Это объясняется универсальностью строения элементарного мышечного волокна (миофибриллы) и тождественностью молекулярного механизма его сокращения. Поскольку гомологичные мышцы в среднем укорачиваются на одну и ту же долю (примерно на 30 %), то чем мышца длиннее, тем на большую длину она сокращается. Следовательно, работа, совершаемая мышцей при сокращении, пропорциональна ее сечению и длине, т. е. пропорциональна ее объему (или массе). Теперь посмотрим, как все это отражается на двигательной активности животного и прежде всего на его беге. Бег состоит из последовательности циклов сокращения и расслабления практически всех скелетных мышц. При каждом таком цикле животное перемещается на один шаг, совершая работу, пропорциональную массе своей мускулатуры. А так как эта последняя у млекопитающих составляет в среднем около 40 % от массы тела, то получается, что работа по перемещению на один шаг пропорциональна массе животного, т. е. Аш~ М. (2.7) Очевидно, что чем длиннее ноги у животного, тем больше его шаги, т. е. тем меньше нужно их сделать для преодоления одной и той же дистанции. Как показывают измерения, у млекопитающих разных видов длина ног пропорциональна M0,33. Значит, число шагов (n), нужное для покрытия фиксированного расстояния, скажем, один километр, должно быть связано с массой тела соотношением n ~ М-0,33. (2.8) При этом животное совершает работу A, равную nAш, т. е., учитывая (2.7) и (2.8), A ~ М0,67. (2.9) Поделив обе половины этого соотношения на М, получим, что работа по транспорту единицы массы тела на фиксированное расстояние (A0) должна уменьшаться при укрупнении животного, точно отражая сокращение необходимого числа шагов: А0 ~ М-0,33. (2.10) Объем работы А0 можно оценить по количеству кислорода, необходимому для переноса килограмма массы тела на один километр. Оказалось, что эта величина ц — кислородная цена транспорта единицы массы — падает с увеличением размеров животного, подчиняясь аллометрическому уравнению ц = c*M-0,33. (2.11) Ясно, что кислородная цена транспорта всей массы животного на один километр (Ц) будет в М раз больше, т. е. Ц = с*М0,67. (2.12) Самая поразительная особенность этого уравнения заключается в том, что оно справедливо для всех бегающих наземных животных — ящериц, млекопитающих и даже насекомых. Значение коэффициента с изменяется у разных групп очень незначительно — от 0,6 до 0,8 литра кислорода для перемещения одного килограмма на один километр. Фактически это означает, что эффективность локомоторного аппарата у всех наземных животных примерно одна и та же, вне зависимости от их эволюционной продвинутости. Скорее всего это постоянство зиждется на единстве строения миофибриллы — основной рабочей структуры любой скелетной мышцы. Теперь посмотрим, как связана с массой тела мощность, развиваемая животным при беге. Напомним, что мощность — это работа, совершаемая за единицу времени. Заменив в уравнении (2.6) величину Δl на абсолютную скорость укорочения мышцы v, мы получим формулу для оценки мощности мышечного сокращения (Pm): Pm = S*σ*v. (2.13) Поскольку максимальные значения v для мышц разных видов животных (при одной температуре) близки, то максимальная мощность мышцы определяется площадью ее поперечного сечения S. Эта площадь при изометричном росте должна увеличиваться пропорционально М0,67, и точно так же должна возрастать мощность мышечного сокращения. Но, как мы знаем (см. уравнение (2.3)), мощность метаболизма увеличивается с ростом массы существенно быстрее. Для метаболизма покоя показатель степени при массе равен 0,75, а при нагрузках он достигает у млекопитающих даже 0,81.
 Рис. 7. Усложнение ветвления одномерных структур при увеличении размеров органов.
а — простая структура; б — сложная.
Рис. 7. Усложнение ветвления одномерных структур при увеличении размеров органов.
а — простая структура; б — сложная.
По-видимому, некоторое превышение реальной мощности метаболизма над уровнем, достаточным для снабжения энергией скелетной мускулатуры, ведет к ускорению бега животного. Скорость бега можно определить, перемножив частоту шагов на длину среднего шага. Измерения показали, что с увеличением размеров животных частота их шагов снижается существенно медленнее, чем возрастает длина шага, в итоге бег ускоряется. Итак, увеличенные размеры дают определенные преимущества. Во-первых, без снижения скорости (и даже при ее некотором росте) бег становится более экономичным. Во-вторых, падает удельная мощность метаболизма, что фактически означает уменьшение энергетических расходов на поддержание жизни единицы биомассы. Однако эти, казалось бы, явные плюсы сменяются на минусы при беге в гору. Для поднятия единицы массы тела на одну и ту же высоту животным вне зависимости от их размеров нужно затратить одно и то же количество энергии, но удельная мощность метаболизма у мелких животных больше, поэтому при беге в гору им надо повысить эту мощность в меньшее число раз, чем крупным животным. Так, при беге по вертикали мышь увеличивает потребление кислорода на 20 %, а лошадь — на 630 %. Значит, бегать по пересеченной местности мелким животным легче.
 Рис. 8. Усложнение поверхности зубов у лошадей в процессе эволюции (по: [Хадорн, Венер, 1989]).
а — Eohippus (эоцен); б — Mesohippus (олигоцен); в — Merychippus (миоцен); г — Equus (плиоцен — современность).
Рис. 8. Усложнение поверхности зубов у лошадей в процессе эволюции (по: [Хадорн, Венер, 1989]).
а — Eohippus (эоцен); б — Mesohippus (олигоцен); в — Merychippus (миоцен); г — Equus (плиоцен — современность).
Время жизни особи принято измерять годами, днями и т. д., хотя его можно было бы отсчитывать и числом каких-нибудь внутренних циклов — числом сердцебиений, дыхательных движений, опорожнений кишечника, делений клеток определенного типа и т. д. На выполнение каждого такого цикла требуется энергия, в связи с этим поток энергии, приходящийся на единицу массы тела (т. е. удельную мощность метаболизма р), можно выбрать в качестве меры скорости протекания всевозможных циклических процессов. Чем эта скорость выше, тем больше циклов совершается в организме за единицу астрономического времени, т. е. тем быстрее течет собственное время индивида. Темп жизни животного ускоряется с уменьшением его массы (см. (2.5)). Это означает, что мелкие животные способны реагировать на изменение окружающей среды быстрее, чем крупные. Обратная сторона медали заключается в ослаблении устойчивости к энергетическому голоду. Хорошо известно, что для крошечных землероек прекращение питания даже на 7–9 часов равносильно гибели. Попробуем выяснить, как соотносятся между собой размеры тела и рассмотренная выше сложность морфологических структур. Сначала отметим, что увеличение органа может привести к усложнению одномерных структур, обеспечивающих связь этого органа с другими частями организма. На рис. 7 изображено, как повышается ранг ветвления сосудов, протоков, нервов и т. д. Кроме того, есть веские основания считать, что изометрический рост ведет к непропорциональному увеличению функциональной нагрузки на многие органы. Мы уже отмечали, что при укрупнении животных их суммарный метаболизм возрастает по меньшей мере пропорционально М0,75. Однако при изометричном росте площади двухмерных структур увеличиваются существенно медленнее, следуя за М0,67. Если эти площади прямо отражают мощности соответствующих структур, то пропорциональный рост приведет к их относительному ослаблению. Отсюда следует, что функционально важные поверхности должны расти быстрее, чем М0,67, т. е. аллометрично. В качестве примера посмотрим, как увеличение размеров животного может отразиться на строении зубов. Пусть в эволюции каких-нибудь травоядных идет увеличение размеров тела, но характер пищи при этом не изменяется. Ясно, что количество пищи, потребляемое особью за сутки, должно соответствовать количеству поглощенного кислорода, т. е. способность перетирать пищу должна соответствовать мощности метаболизма. Поэтому при увеличении массы особи рабочая поверхность зубов должна расти быстрее, чем поверхность тела. Данное требование удовлетворяется за счет появления на перетирающей поверхности зубов характерных выпячиваний — эмалиевых складок и бугорков. Тенденция к усложнению поверхности зубов при увеличении размеров животных хорошо прослеживается в эволюционных рядах лошадей и других травоядных (рис. 8). Иллюзорность физиологического прогресса Под общественным прогрессом мы обычно понимаем исторический процесс постепенного устранения препятствий к исполнению желаний каждого человека, обусловленных его биологической и социальной природой. Высшая стадия исторического развития представляется нам обществом материального изобилия и полной социальной справедливости, где «все равны» и «каждому по потребностям». Обычно человек стремится в своей жизни к двум целям: во-первых, застраховать себя и свою семью от теоретически возможной угрозы голода и холода (что достигается накоплением некоторого избытка энергии «на черный день») и, во-вторых, занять более высокое место в обществе. В итоге в ходе истории человечества четко прослеживаются две тенденции — рост потребления энергии на душу населения и постепенное выравнивание социальных рангов, т. е. демократизация человеческого общества. Если общественная жизнь людей отличается многими видоспецифичными особенностями, то стремление к независимости от капризов среды разделяют с нами все живые существа. По-видимому, именно поэтому мы склонны считать, что эволюционный прогресс организмов тесно связан с повышением способности потреблять с большей эффективностью возрастающее количество энергии. Впервые такое представление о прогрессивной эволюции было сформулировано А.Н. Северцовым, который назвал ее морфофизиологическим прогрессом, или ароморфозом. Данная форма эволюции характеризуется повышением степени морфологической дифференциации организма и интенсификацией функций, что ведет к подъему общего уровня «энергии жизнедеятельности». Эта точка зрения на эволюционный прогресс разделяется большинством советских биологов. Наш крупнейший эволюционист И.И. Шмальгаузен видел в ароморфозах «процессы эволюции, освобождающие организмы от слишком тесных ограничений в связях со средой и как бы подымающие их над многими частными условиями». Если под несколько неопределенным выражением «подъем энергии жизнедеятельности» понимать повышение мощности метаболизма, то получается, что скорость, с которой организм извлекает из внешней среды энергию и вещество, возвращая обратно тепло и отбросы, является мерой его эволюционной продвинутости. И.И. Шмальгаузен считал, что организм «получает в борьбе за существование преимущества общего характера, не ограниченные какой-либо строго определенной средой, и поэтому приобретает возможность выйти за пределы той среды, в которой жили его предки, и захватить новые, частью весьма отличные области обитания». Классическим примером ароморфоза является приобретение предками птиц и млекопитающих теплокровности. Повышение температуры тела на 10 °C ведет к ускорению всех химических реакций в 2–3 раза, что вроде бы дает организму явные преимущества. Достаточно обратить внимание на то, что теплокровные животные сохраняют активность при пониженных температурах, когда их холоднокровные собратья впадают в оцепенение. Таким образом, приобретение теплокровности (во всяком случае на первом этапе) резко повышает возможность добывания пищи при сокращении числа потенциальных хищников и конкурентов. Мощность метаболизма у млекопитающих примерно в 10 раз выше, чем у амфибий и рептилий той же массы. Резко возросший поток кислорода требует пропорционального увеличения рабочей поверхности легких. Один кубический сантиметр легочной ткани лягушки имеет газообменную поверхность 20, а человека — 300 квадратных сантиметров. Повышение метаболизма особи предполагает пропорциональное увеличение пищевого потока, для чего необходимо укрупнение и усложнение многих рабочих структур, обслуживающих функции доставки, удаления и гомеостаза. Среди структур, принимающих участие в обеспечении пищевого потока, немалая роль принадлежит центральной нервной системе. Для наземных позвоночных одного класса масса головного мозга m связана с массой тела М аллометрическим уравнением т = b*М0,67. (2.14) Прогрессивное развитие головного мозга у млекопитающих по сравнению с рептилиями выражается в многократном увеличении коэффициента b уравнения (2.14): от 0,001 (типичного для рептилий) до 0,01 (среднего для млекопитающих). Между прочим, коэффициент b хорошо передает прогрессивное развитие мозга в линии, ведущей к человеку. Для обезьян (в целом) он равен 0,02—0,03, для антропоидов — 0,03—0,04, для человека —0,08—0,09. Этот восходящий ряд и является «объективным» основанием для нашего антропоцентризма. Суммируя сказанное, можно считать, что переход к теплокровности означал существенное усложнение большинства рабочих структур, обслуживающих по меньшей мере три из пяти основных функций. Этот сдвиг вполне можно назвать повышением организации. Однако задумаемся, чему служат все эти многочисленные структуры? Казалось бы, прежде всего тому, чтобы обеспечить организм обильным питанием. Действительно, млекопитающие и птицы (по сравнению с амфибиями и рептилиями) из-за повышенного темпа жизни могут добыть себе за единицу времени гораздо больше пищи, но, увы, они по той же причине и должны потреблять ее гораздо больше, а голодание переносится ими много хуже. Тогда о каком же уменьшении зависимости от среды может идти речь? Ясно, что повышенная мощность пищедобывающих структур является в данном случае суровой необходимостью. Скрупулезное изучение степени развития органов и структур у млекопитающих привело американских физиологов К.Тэйлора и Э.Вейбеля к важнейшему эмпирическому обобщению, названному ими принципом симморфоза. Этот принцип гласит, что мощность ни одной рабочей структуры в организме не превосходит уровня, необходимого животному при максимальных нагрузках. Короче говоря, в организме нет ничего избыточного. По выражению крупнейшего специалиста по сравнительной физиологии К. Шмидта-Ниельсена, «каждый организм можно рассматривать как оптимальную для своих размеров и типа устройства конструкцию». Как же возникла такая поразительная целесообразность? Очень просто, из-за регулярной нехватки пищевых ресурсов. Гонка и голод — вот причина симморфоза. Принцип симморфоза заставляет сомневаться в каком-то организационном преимуществе одних форм над другими, так как все они сидят на очень скудном, полуголодном пайке и неспособны выкроить средства на роскошь обладания сверхразвитыми структурами. Теперь поговорим о «победе», одержанной теплокровными. Заметим, что каждая особь теплокровных животных нуждается в большей территории, чем особь той же массы у рептилий и амфибий. Значит, плотность популяций теплокровных животных должна быть в среднем ниже, а это отнюдь не способствует выживанию при колебаниях пищевых ресурсов. В настоящее время на Земле существует 2,1 тысячи видов земноводных, около 6 тысяч рептилий, из которых 5,7 тысячи ящериц и змей, около 8 тысяч птиц и около 4 тысяч млекопитающих. Если скинуть со счетов птиц (как обитателей особой, воздушной, среды), то соотношение видового многообразия холоднокровных и теплокровных складывается не в пользу последних. И это-то после массовых вымираний тысяч видов динозавров и прочих завров! Выше мы допустили, что особенно ярко преимущество теплокровности должно было проявиться сразу после ее возникновения, когда первых зверей окружали более низкоорганизованные рептилии. Потом же захватившие господство млекопитающие стали конкурировать друг с другом, что называется, на равных. Однако палеонтология свидетельствует, что звери с юры до середины третичного периода (т. е. на протяжении более 100 миллионов лет) были представлены мелкими, величиной с крысу, формами. И, самое главное, их общая видовая численность была ничтожной по сравнению с современной. Мощная адаптивная радиация млекопитающих началась намного позже. Создается впечатление, что их видообразованию сильно мешали более «примитивные» динозавры и другие крупные рептилии. Энергия и скорость Прежде всего отметим, что под эволюционным прогрессом традиционно понимают эволюционную тенденцию к повышению организации живых существ. Можно выделить два подхода к этой проблеме — структурный и функциональный. Если считать организм совокупностью его органов, то повышение организации можно свести к увеличению числа и многообразия функционально активных элементов, составляющих органы. В то же время при сопоставлении эволюционно разобщенных организмов принято обращать внимание на степень интеграции их частей в единое целое. При этом основной упор нередко делается на уровни развития структур, осуществляющих нейрогуморальную регуляцию. Тем самым прогресс организмов сводится к прогрессу некоторых органов, признанных по тем или иным соображениям ведущими. При функциональном подходе внимание фиксируется на взаимодействии организмов с окружающей средой. Здесь можно выделить две стороны проблемы, два ее аспекта — энергетический и динамический, их также можно было бы назвать экстенсивным и интенсивным. Первый основан на оценке суммарной мощности метаболизма индивида, второй — на оценке удельной мощности метаболизма (р), которая показывает, с какой скоростью поглощает энергию среды каждая единица биомассы популяции, т. е. величина р отражает динамизм энергетических потоков, связывающих вид с его средой. Прогрессивное развитие органов неразрывно связано с увеличением их рабочей мощности, поэтому повышение организации обычно выражается в усилении энергетических потоков как внутри организма, так и между организмом и средой. Вместе с тем, мы считаем прогрессивными все изменения, которые ведут к возрастанию скорости, с которой каждая особь популяции реагирует на изменения среды. Обычно эта скорость ассоциируется с усложнением центральной нервной системы, хотя нет оснований умалять в этом отношении другие органы (например, локомоторный аппарат). Интегральной характеристикой скорости реагирования (т. е. мерой динамического прогресса) мы выбрали удельную мощность метаболизма. Соотношение (2.5) показывает, что данный параметр (в какой-то степени отражающий интенсивность метаболизма усредненной клетки) снижается при укрупнении особей, сопровождая процессы усложнения органов и усиления суммарного энергетического потока между организмом и средой. Это сталкивает нас с очевидным парадоксом: динамический прогресс нередко сочетается с энергетическим регрессом и наоборот. Получается, что идти сразу по двум сторонам дороги прогресса не так-то просто. Между прочим, некоторые эволюционные завоевания можно расценить как попытку осуществить одним приемом обе формы эволюционного прогресса. Например, появление теплокровности повышает скорость реагирования популяции на изменения среды при усилении энергетического потока, поглощаемого средней особью. Отметим, что если польза динамического прогресса в «борьбе за жизнь» более или менее очевидна, то рациональный смысл энергетического прогресса от нас постоянноускользает. Неужели причина глобальной тенденции к усложнению структур кроется в прозаичной экономии ресурсов, приходящихся на один грамм биомассы? Действительно, на энергетический прогресс можно взглянуть как на тенденцию к сокращению числа «ртов», «кормящих» биомассу популяции. Если количество пищи, проходящее через каждый рот, при этом растет, то потребность в ней каждой клетки индивида (а стало быть, и всей популяции с фиксированной биомассой) падает. Получается, что энергетический прогресс ведет к более эффективному преобразованию энергии и вещества окружающей среды в биомассу вида. Итак, что же выбрать критерием прогресса? Если удельную мощность метаболизма, то следует признать самыми продвинутыми воробьиных птиц. Если сложность строения органов, то более прогрессивными можно назвать крупных теплокровных животных. Достаточно вспомнить об очень сложных зубах крупных травоядных, сложном желудке крупных жвачных, переднем мозге с большим числом извилин высших приматов и китообразных. Однако не следует забывать, что идущее при укрупнении особей ослабление клеточного метаболизма означает снижение скорости многих синтетических процессов. Какая клетка совершеннее — с высоким или низким метаболизмом? Скорее всего с высоким. Ведь чем выше уровень клеточного метаболизма, тем выше скорость всех циклических процессов и тем лучше «подогнаны» друг к другу активности многих генов. Вполне очевидно, что такое состояние менее случайное, т. е. оно соответствует более высокому уровню генетической информации. Таким образом, усложнение на надклеточном уровне как бы компенсируется некоторым упрощением на субклеточном и молекулярном уровнях.
Микроэволюция
Глава 3. Факторы микроэволюции
В природе существует много того, что не может быть ни достаточно глубоко понято, ни достаточно убедительно доказано, ни достаточно умело и надежно использовано на практике без помощи и вмешательства математики.Для того чтобы в вопросах эволюции оставаться на твердой почве фактов, разумно руководствоваться принципом униформизма (актуализма), т. е. при объяснении явлений отдаленного прошлого исходить из анализа процессов, совершающихся в настоящее время. Не будем спешить и придумывать особые законы, влекущие эволюцию организмов по пути морфофизиологического прогресса. Сначала постараемся понять протекающую перед нашими глазами жизнь природных популяций, после чего сделаем попытку распространить эти (микроэволюционные) процессы на гораздо большие отрезки времени. Генный поток Сначала попробуем ответить на вопрос: что такое вид? Это понятие относится к наиболее важным и сложным в современной биологии. Начнем с того, что особи, связанные отношением родства, принадлежат одному виду. Под родством же люди обычно подразумевают каким-то образом зарегистрированную общность происхождения. Например, братья и сестры имеют общими двух предков в поколении родителей. Двоюродные братья и сестры имеют общих предков в поколении дедов, троюродные — в поколении прадедов и т. д. Таким образом, с понижением степени родства общие предки перемещаются на все более отдаленные поколения. Совокупность особей, связанную отношением родства (включая и самые отдаленные его степени), можно было бы назвать видом. По существу, это очень близко к библейскому определению рода человеческого как всех потомков Адама и Евы. Однако такое определение крайне неудобно и ненадежно. Разве особи разных видов (например, волки и лисицы) не связаны отдаленным (эволюционным) родством? Как определить ту степень родства, которая еще позволяет отнести организмы к одному виду? С генетической точки зрения, от предка к потомку передаются практически в неизменном виде наследственные факторы, гены. Поэтому общность происхождения — это общность генов. Миф об Адаме и Еве на самом деле довольно точно отражает реальную ситуацию, во всяком случае, есть веские основания считать, что вся совокупность наследственных факторов человечества восходит к генам очень небольшой группы особей. И если бы гены при передаче от родителей к потомкам не изменялись (т. е. не мутировали), то все люди были бы действительно братьями. В ядре любой соматической (т. е. не половой) клетки организма имеются два аллельных варианта каждого гена, два аллеля — один от матери, другой от отца. Исключение составляют только половые клетки — гаметы, в их геноме каждый локус представлен всего одним аллелем, такой набор генов называется гаплоидным. При оплодотворении гаплоидные геномы гамет соединяются в диплоидный геном зиготы, который затем точно копируется механизмом клеточного деления. Таким образом, геномы всех соматических клеток организма на всех стадиях его развития (включая и зиготу) идентичны. Гаплоидный геном гамет возникает в результате действия особого клеточного механизма — мейоза. Суть его состоит в том, что в одну гамету попадает только один аллельный вариант каждого генного локуса, какой именно — материнский или отцовский — решает случай. Сделаем одно важное (и в какой-то степени спорное) предположение. Будем считать, что все особи одного вида обладают набором одних и тех же генных локусов, т. е. при мутировании может изменяться не число, а только аллельное состояние локусов. Потерю гена формально можно трактовать как его мутирование к нулевому аллелю. Правда, с добавлением гена (его дупликацией) вопрос обстоит сложнее. Итак, примем, что все особи одного вида содержат в хромосомах каждой клетки (в своем геноме) одно и то же число генных локусов, или генов. Поскольку при передаче наследственного материала от родителя к потомку с некоторой вероятностью может произойти мутация по любому гену, то чем ниже степень родства особей, тем выше вероятность различия аллелей по каждому локусу. Различие аллелей ведет к различию фенотипов, т. е. к уменьшению внешнего сходства. Следовательно, степень родства можно оценить по близости внешнего вида особей. Мы оцениваем такое сходство на глаз, интуитивно схватывая огромное число признаков. В итоге, глядя на окружающие нас живые организмы, мы можем рассортировать их по внешнему виду, что, как правило, эквивалентно разбиению по видовой принадлежности. Нет ли способа объективно оценивать степень сходства генов? Оказывается, такой способ есть, и им уже давно пользуются биологи. Считается, что особи разного пола принадлежат к одному виду, если они способны к самостоятельному скрещиванию и их потомство плодовито. При оплодотворении две гаметы родителей сливаются в зиготу, при этом объединяются два набора аллелей — от отца и матери. Под контролем конкретной комбинации аллелей (генотипа) зигота превращается во взрослый организм. В связи с этим можно говорить о генетической программе развития организма, хотя это выражение не более, чем метафора, во всяком случае данная «программа» не имеет ничего общего с компьютерной. Итак, онтогенез потомка протекает под контролем сразу двух генетических программ. Если они не очень хорошо согласованы, то онтогенез может либо вовсе не завершиться, либо привести к созданию дефектной особи. Таким образом, метод скрещивания фактически оценивает близость аллелей по локусам, контролирующим онтогенез. Именно этим методом и пользуются особи одного вида в своей естественной жизни. Впрочем, для многих животных со сложным поведением оценка видовой принадлежности по внешним признакам также является необходимым условием для скрещивания, и спаривания не произойдет, если особи разных полов не «поймут» ритуала ухаживания. Ситуация, когда любые две взрослые разнополые особи одного вида могут стать участниками скрещивания, называется панмиксией. Естественно, в реальности условия для панмиксии выполняются далеко не всегда, поскольку для соединения любой пары гамет нужно, Чтобы родители — обладатели гамет — находились недалеко друг от друга. Тем не менее в пределах этого «недалеко» панмиксия выполняется. Например, жителей большого этнически однородного города с хорошо организованным транспортом можно считать связанными панмиксией. У большинства зоологических видов особи способны к активному движению, кроме того, могут двигаться их личинки или гаметы. У ботанических видов к широкому расселению способны семена, споры, пыльца, а нередко и фрагменты растений. Мы можем взглянуть на это явление немного иначе и увидеть в нем блуждание генов по ареалу вида. Данный процесс имеет много общего с диффузией броуновских частиц. Поток таких частиц в каком-то направлении пропорционален градиенту (перепаду) их концентрации вдоль этого направления и коэффициенту диффузии — параметру, отражающему подвижность самих частиц. В конечном счете концентрация частиц во всей области их блуждания выравнивается, и наступает диффузионное равновесие. Точно так же по ареалу вида «движутся» и аллели. Как и в случае диффузии, через некоторое время, определяемое размерами ареала вида и способностью генов к перемещению, должно установиться равновесие аллельных частот. Если скорость движения генов высока, то процесс выравнивания этих частот идет быстро. При панмиксии, когда в скрещивание может вступить любая пара разнополых особей, аллельное равновесие устанавливается практически моментально, т. е. за одно поколение. Нередко мы имеем дело только с переходом к такому равновесию, когда на территории, занятой видом, обнаруживаются градиенты по частотам разных аллелей. Эти градиенты — клины — можно изобразить графически в виде семейства изолиний, как это принято для топографических карт. Вдоль такой изолинии частота аллеля постоянна. Где изолинии гуще, градиент частоты аллеля круче. По аналогии с диффузионным потоком можно ввести понятие о генном потоке как о переносе аллелей через единицу длины изолинии. Ясно, что такой поток будет тем мощнее, чем круче градиент и чем больше коэффициент диффузии генов. Наиболее мощные генные потоки образуются, когда в какую-то область ареала с высокой концентрацией определенного аллеля проникает группа иммигрантов с другой частотой того же аллеля. Представим себе, что область блуждания броуновских частиц разделена на несколько отсеков перегородками с разной проницаемостью. Понятно, что если все частицы поместить в один из таких отсеков, то характер последующей диффузии будет существенно зависеть от геометрии расположения перегородок и проницаемости каждой из них. Точно так же территория, где обитают особи одного вида, далека от однородности. Здесь могут быть и реки, и горы, и луга, и леса, — и все это создает препятствия (барьеры) для генных потоков. Ясно, что если вид занимает сильно пересеченную местность, то время выравнивания концентраций аллелей будет много больше, чем в случае однородного ландшафта. Более того, в реальной ситуации существует немало районов, отделенных от остальных частей ареала практически непроницаемыми барьерами. Представим ареал как замкнутое пространство, разделенное на ячейки барьерами разной проницаемости. Внутри таких ячеек препятствий для генного потока практически нет, поэтому аллельные частоты в них выравниваются сравнительно быстро. Во всяком случае, скорость этого процесса внутри ячеек на несколько порядков выше, чем на их границе. Ясно, что даже в соседних ячейках частоты одних и тех же аллелей могут резко различаться. Вот эту-то часть вида, населяющую одну ячейку, и назовем популяцией. Каждая популяция может быть охарактеризована своим генофондом, т. е. составом аллелей каждого локуса. Популяцию, занимающую ячейку с непроницаемыми стенками, назовем изолированной.Ф.Бэкон
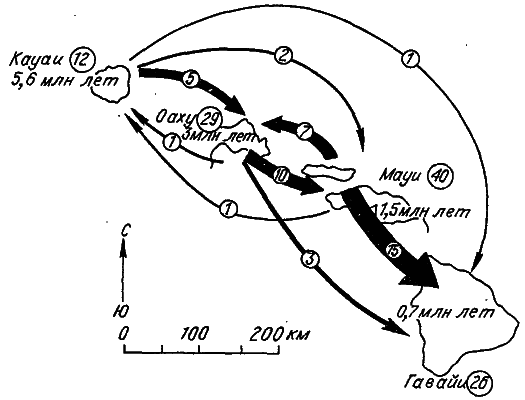 Рис. 9. Схема видообразования гавайских дрозофилид (по: [Carson, 1983]).
Стрелками показаны направление и число заносов мух-основательниц новых видов. В кружках — количество видов-эндемиков каждого острова.
Рис. 9. Схема видообразования гавайских дрозофилид (по: [Carson, 1983]).
Стрелками показаны направление и число заносов мух-основательниц новых видов. В кружках — количество видов-эндемиков каждого острова.
Встает естественный вопрос, как особи попадают в изолированные ячейки? Можно лишь заметить, что абсолютно непроницаемых барьеров нет, кроме того, их проницаемость со временем изменяется. Если виды существуют в течение миллионов лет, то легко себе представить, что большинство барьеров в их ареалах изменяют за это время свою проницаемость в широчайшем диапазоне. Даже такой молодой вид, как Н. sapiens (его возраст оценивают всего в 40 тысяч лет), уже успел столкнуться с неоднократным наступлением и отступлением ледника, представляющего собой весьма эффективный изолирующий барьер. Американский генетик Х. Карсон сумел восстановить родословную одной группы дрозофилид, обитающих на Гавайских островах (рис. 9). Эти вулканические острова возникали друг за другом в направлении с северо-запада на юго-восток. Самому древнему острову Кауаи 5,6 миллиона лет, а самому молодому острову Гавайи — всего 700 тысяч лет. Анализ политенных хромосом 97 видов гавайских дрозофил привел Карсона к заключению, что все они происходят от одного вида, обитавшего на острове Кауаи. Затем мухи проникли на более юный остров Оаху и, наконец, на остров Гавайи. Число родственных видов во много раз превосходит число островов, поэтому следует предположить, что иногда происходило попадание мух с одного заселенного острова на другой. При этом, несмотря на общность происхождения, иммигранты не скрещивались с туземцами, т. е. обе группы дрозофилид за время географического разобщения успевали стать разными видами. Залеты мух с острова на остров относятся к очень редким событиям, поэтому скорее всего в основе каждого нового вида стояла всего одна осемененная самка. Этот пример показывает, что для интеграции генофондов популяций в единый генофонд вида продолжительность существования барьеров должна быть намного меньше времени превращения изолированной популяции в новый вид. Очевидно, что со временем в разделенных друг от друга частях некогда единой популяции могут произойти изменения аллельного состава генов, достаточные для того, чтобы воспрепятствовать спариванию особей или нарушить нормальное течение онтогенеза их потомков; иными словами, разобщенные генофонды станут несовместимыми. Такая несовместимость представляет собой особый, самый эффективный, генетический барьер, разделяющий популяции навсегда. Итак, на популяции одного вида постоянно действуют два фактора, противоположных по своему конечному эффекту. Генный поток стремится слить генофонды популяций в единый генофонд, тогда как процесс видообразования пытается воздвигнуть между временно разобщенными популяциями непреодолимые генетические барьеры. Далее нам предстоит понять, какие факторы лежат в основе генетической дифференциации популяций. Элементарный акт микроэволюции Очевидно, что в ходе эволюции идет замещение одних аллелей на другие в ряде локусов. Будем считать элементарным микроэволюционным актом замещение в одном из локусов старого аллеля на новый у всех особей популяции. Учитывая огромные численности природных популяций (105 — 109 особей), «элементарный акт» микроэволюции требует для своего завершения длительного времени. В связи с этим вспомним некоторые рассмотренные выше эволюционные теории. С точки зрения приверженцев номогенеза, эволюционный сдвиг развивается под влиянием каких-то внутренних факторов без всякой связи с изменениями окружающей среды. По мнению Л.С. Берга, «при возникновении новых географических форм (видов, подвидов, наций) образованием новых признаков захватывается громадная масса особей, обитающих в определенной географической местности». Такое явление можно было бы объяснить, допустив строгую направленность характера мутирования, когда у большинства особей популяции в клетках зародышевого пути в значительной части локусов происходит превращение старых аллелей во вполне конкретные новые. Фантастичность такого представления самоочевидна. С точки зрения ламаркизма, процесс возникновения новых аллелей уже с самого начала имеет приспособительное значение. Иными словами, между изменениями среды и характером мутирования должна существовать связь, обусловленная каким-то неизвестным молекулярным механизмом. Главное состоит в том, что возникшие мутанты должны быть лучше приспособлены к окружающей среде. Скажем, если в среде обитания популяции падает концентрация кислорода, то в локусах, кодирующих полипептиды гемоглобина, возникают мутации, повышающие сродство этого белка к кислороду. Если же затем в окружающей среде концентрация кислорода вновь повышается, то локусы гемоглобина мутируют в обратном направлении. Заметим, что эти мутации должны происходить в клетках зародышевого пути, в которых гены гемоглобина даже не экспрессируются. Все наши знания о характере мутирования не соответствуют такому ходу событий. Наиболее спокойной, целиком основанной на известных фактах, представляется нам точка зрения неодарвинизма (вейсманизма). Новые аллели в данном случае возникают совершенно спонтанно: ни скорость возникновения мутаций, ни их эффект никак не связаны с изменениями окружающей среды. При этом каждый локус можно охарактеризовать особым свойством— мутабильностью, т. е. вероятностью смены им аллельного состояния. Для ряда генов, изменения которых ведут к четким, легко регистрируемым изменениям фенотипа особей, мутабильность была измерена. Для совершенно разных организмов она составила 10-5—10-6 на одну копию локуса за генерацию. Иными словами, из каждых 100 тысяч особей, возникших в популяции за поколение, лишь одна может оказаться носителем новой мутации по конкретному гену. Заметим, что для завершения элементарного микроэволюционного акта мутантный аллель должен заменить старые аллели во всех экземплярах этого локуса в популяции. Расчеты показывают, что надеяться исключительно на процесс мутирования по меньшей мере неразумно. Даже при строгой направленности перехода A1 → А2 (где А2 — новый аллель, а А1 — старый) для превращения только половины старых аллелей в новые при мутабильности 10-5 надо ждать около 70 тысяч поколений. К тому же старый аллель может превратиться в совсем другие аллели — А3, А4 и т. д., и их эффект на фенотип совсем не обязан совпадать с эффектом аллеля А2. Если представить, например, что доля новых аллелей, изменяющих фенотип особи в одном направлении, равна доле аллелей с противоположным эффектом, то средний фенотип особи популяции вообще останется неизменным, лишь постепенно будет возрастать многообразие аллелей, а следовательно, и многообразие фенотипов. Итак, с неодарвинистской точки зрения, элементарный микроэволюционный акт не может быть обеспечен одним мутированием, для этого нужны дополнительные факторы. Естественный отбор в популяциях Мы уже упоминали о том, что Ч. Дарвин ввел понятие естественного (природного) отбора по аналогии с отбором искусственным. Последний сознательно или бессознательно осуществляется человеком, который отбирает на потомство особей с «полезными» для него свойствами. При естественном отборе в качестве селекционера выступает сама природа. Но сказать «природа», значит, не сказать ничего, с равным успехом можно было бы употребить и слово «бог». Какие же реальные факторы стоят за словами «естественный отбор»? Сам Дарвин трактует это понятие следующим образом: «Я назвал это начало, в силу которого каждое незначительное изменение, если оно полезно, сохраняется, естественным отбором…» Иными словами, природа находит «полезным» любое наследуемое свойство особей, которое повышает их жизнеспособность, т. е. их шансы на выживание в борьбе за жизнь. Попробуем подойти к этому немного строже. Определим жизнеспособность как вероятность особи завершить свой онтогенез. Раз дарвиновская «полезность» наследуется, значит, можно найти гены, чьи аллели обеспечивают своих носителей разной жизнеспособностью. В популяционной генетике каждому генотипу приписывается его приспособленность — величина, которая показывает, каковы у обладателей данного генотипа шансы на вклад своих аллелей в генофонд взрослых потомков. Легко заметить, что понятие «приспособленность» шире, чем «жизнеспособность». Ведь особи с равной выживаемостью могут различаться по числу производимых гамет, шансам найти партнера по спариванию, оплодотворяющей способности гамет и т. д. Очевидно, что все такие наследуемые свойства дают их носителям больше шансов передать свои гены потомкам. В связи с этим выделяют несколько компонент приспособленности — жизнеспособность, плодовитость, скорость индивидуального развития и др. В рассматриваемой ниже простейшей модели отбора приспособленность отождествляется только с одной из ее главных компонент — жизнеспособностью. Пусть в популяции, насчитывающей N особей, локус А представлен всего двумя аллелями А1 и А2. Тогда особи должны разделиться на три генотипических класса: 1) гомозиготы А1А1 с численностью N11; 2) гомозиготы А2А2 с численностью N22; 3) гетерозиготы А1А2 с численностью N12, т. е. N = N11 + N22 + N12. (3.1) Каждая особь вносит в генофонд популяции два экземпляра гена А, или (выражаясь несколько иначе) каждая особь является донором (или поставщиком) двух экземпляров этого локуса. Гомозиготы А1А1 вносят 2N11 копий аллеля А1, гомозиготы А2А2 — 2N22 копий аллеля А2, тогда как гетерозиготы А1А2 — 2N12 копий аллеля А1 и столько же копий аллеля А2. Отсюда легко оценить долю (частоту) каждого аллеля среди всех 2N экземпляров локуса А, которыми располагает генофонд популяции. Обозначим частоту аллеля А1 через р, а частоту аллеля А2 через q (отметим, что р + q = 1). Эти частоты можно вычислить по формулам p = (2N11 + N12) / 2N = (N11 + 0,5N12) / N, (3.2) q = (2N22 + N12) / 2N = (N22 + 0,5N12) / N. (3.3) Гетерозиготы являются донорами сразу двух аллелей, в связи с этим (чисто формально) можно считать, что одна половина гетерозигот вносит в генофонд аллель А1, а другая — аллель А2. Тогда всех особей популяции можно представить в виде суммы двух классов — доноров аллеля А1 с численностью N1 и доноров аллеля А2 с численностью N2. При этом N1 = N11 + 0,5N12, (3.4) N2 = N22 + 0,5N12. (3.5) Взглянув на правые части равенств (3.2) и (3.3), мы видим, что в популяции частота каждого аллеля равна доле его доноров: p = N1 / N; q = N2 / N. (3.6) Рассмотрим популяцию на двух стадиях развития ее особей — на стадии зиготы и взрослого организма. Обозначим численность зигот через N, а численность взрослых — через N'. Каждая из этих величин (N и N') складывается из численностей трех генотипических классов: N11, N22, N12 — для зигот и N11', N22', N12' — для взрослых. Теперь каждому генотипическому классу припишем его приспособленность: w11 — гомозиготам A1A1, w22 — гомозиготам А2А2 и w12 — гетерозиготам. Определим приспособленность каждого генотипа как долю зигот (с данным генотипом), достигших взрослой стадии, т. е. w11 = N'11 / N11; w12 = N'12 / N12; w22 = N'22 / N22. (3.7) Так же можно ввести представление и о приспособленности любой группы генотипов. Например, можно говорить о средней приспособленности популяции (w) как о доле всех зигот, успешно завершивших развитие. Аналогично определяется средняя приспособленность доноров каждого аллеля — w1 и w2: w = N' / N; w1 = N1' / N1; w2 = N2'/ N2. (3.8) Наша главная задача состоит в том, чтобы определить, как под действием отбора изменяется аллельный состав популяции, иначе говоря, как изменяется частота аллелей А1 и А2 за время онтогенеза особей. Доля аллеля в генофонде популяции равна доле его доноров (формулы (3.6)). В случае аллеля А2 это (N2 / N) — на стадии зигот и (N2' / N') — на стадии взрослых. Поэтому сдвиг частоты аллеля А2 в результате отбора (Δq) можно получить как разность: (N2' / N') — (N2 / N). Выразив численности взрослых через численности зигот (см. уравнения (3.8)), получим Δq = N2*w2 / Nw — N2 / N = q((w2 — w) / w). (3.9) Итак, сдвиг частоты аллеля в генофонде популяции в результате отбора равен частоте этого аллеля до отбора, умноженной на относительное превосходство приспособленности его доноров над средней приспособленностью популяции. Отбор в панмиктической популяции Перейдем к панмиктической популяции. Здесь при оплодотворении аллели гамет соединяются в генотипы зигот по правилам свободного попарного комбинирования. Зиготы с генотипами А1А1, А1А2 и возникают в соотношениях р2: 2pq: q2 (рис. 10). Эти же соотношения, получившие название закона Харди-Вайнберга, будут справедливы и для взрослых членов популяции, если приспособленность всех генотипов одна и та же. Закон Харди-Вайнберга довольно часто выполняется в природных популяциях, он позволяет легко вычислять частоты генотипов, исходя из частот аллелей. В нашей модели отличия в приспособленности особей обусловлены только разным уровнем их жизнеспособности, поэтому численности зигот трех генотипических классов А1А1, А1А2 и А2А2 соответственно равны Np2, 2Npq и Nq2. Тем самым зиготы в точности отражают генофонд популяции взрослых особей предыдущего поколения. В результате действия отбора численности генотипов взрослых особей, развившихся из зигот, приобретают вид N11' = Np2w11; N12' = 2Npqw12; N22' = Nq2w22. (3.10)
 Рис. 10. Схема формирования генотипов зигот при случайном соединении гамет (при панмиксии).
Объяснения в тексте.
Рис. 10. Схема формирования генотипов зигот при случайном соединении гамет (при панмиксии).
Объяснения в тексте.
Численность всех взрослых особей после отбора связана с численностью зигот уравнением N' = N(р2w11 + 2pqw12 + q2w22). (3.11) Поскольку отношение N'/N выражает среднюю приспособленность популяции (см. уравнения (3.8)), поделив обе части уравнения (3.11) на N, получим w = р2w11 + 2pqw12 + q2w22. (3.12) По аналогии с формулой (3.5) можно получить численность доноров аллеля А2 после отбора, т. е. среди взрослых потомков N2' = N22' + 0,5N12', (3.13) или, учитывая соотношения (3.10), N2' = Nq2w22 + Npqw12. (3.14) Поделив обе части этого уравнения на N2 = Nq (см. (3.6)), получим значение приспособленности доноров аллеля А2: w2 = N2' / N2 = (Nq(qw22 + pw12)) / Nq, (3.15) т. е. w2 = pw22 + qw12. Точно так же можно показать, что w1 = pw11 + qw12. (3.16) Итак, мы располагаем всеми данными для вычисления сдвига частот аллелей после отбора (см. уравнение (3.9)). Отбор в смысле Ч. Дарвина основан не на абсолютном уровне истребления молодых особей, а на различиях в степени такого истребления. В связи с этим приспособленность какой-то группы особей удобно объявить равной единице, а приспособленность других групп представить в виде суммы 1 + s (s — коэффициент отбора). Такой прием позволяет рассмотреть наиболее интересную для нас задачу — судьбу мутаций, влияющих на относительную приспособленность их носителей. Судьба вредной мутации Предположим, что в огромной панмиктической популяции возник и каким-то образом распространился новый аллель А2 локуса А. (Ранее этот локус был представлен аллелем А1.) Ясно, что мутантный аллель может встречаться в гомо- и гетерозиготном состояниях. Если мутация вредная, значит, приспособленность гомозигот А2А2 должна быть ниже, чем у гомозигот по нормальному аллелю А1А1. Это обстоятельство мы можем отразить равенствами w11 = 1, w22 = 1—s. Возникает вопрос: какая приспособленность будет у гетерозигот А1А2? В классической генетике рассматриваются три случая: 1) мутация рецессивна, т. е. приспособленность гетерозигот равна приспособленности гомозигот по нормальному аллелю w12 = w11 = 1; 2) мутация доминантна, т. е. приспособленность гетерозигот равна приспособленности гомозигот по мутантному аллелю w22 = w12 = 1—s; 3) случай неполного доминирования, когда приспособленность гетерозигот ниже, чем у гомозигот по нормальному аллелю, но выше, чем у гомозигот по мутантному. Степень доминирования можно оценить с помощью специальной величины А, варьирующей от нуля до единицы. При этом приспособленность гетерозигот можно выразить формулой w12 = 1 — hs, охватывающей все три рассмотренных выше случая. При h = 0 мы имеем дело с рецессивной вредной мутацией, при h = 1 — с доминантной, а при неполном доминировании — 0 < h < 1. Особый интерес для нас представляет случай полудоминирования, когда h = 0,5. Дело в том, что, по-видимому, большинство вредных мутаций, постоянно возникающих в природных популяциях, полудоминантны. Приспособленности генотипов в этом случае определяются соотношениями w11 = 1; w22 = 1 — s; w12 = 1 — 0,5s. (3.17) Эти равенства позволяют вычислять (с помощью формулы (3.12)) значение средней приспособленности популяции: w = р2 + 2pq(1 — 0,5s) + q2(1 — s), или w = (р2 + 2pq + q2) — qs(р + q). Напомним, что p + q = 1, поэтому w = 1 — qs. (3.18) Аналогично с помощью уравнения (3.15) можно оценить приспособленность доноров мутантного аллеля: w2 = q(1 — s) + р(1 — 0,5s) = 1 — qs — 0,5ps. (3.19) Теперь, подставив в уравнение (3.9) только что полученные значения для w и w2, получим сдвиг в частоте мутантного аллеля за одно поколение отбора: Δq = -0,5(pqs / 1 — qs). (3.20) Так как мутация возникла недавно, то ее частота должна быть очень низкой. Если к тому же невелик и коэффициент отбора, то знаменатель правой части уравнения (3.20) мало отличается от единицы. Поэтому сдвиг частоты мутантного аллеля за одно поколение отбора можно оценить с помощью приближенной формулы Δq ≈ -0,5pqs. (3.21) Знак минус в правой части равенства указывает на то, что естественный отбор «стремится» снизить частоту аллеля A2. Эффективность этого отбора, получившего название очищающего, тем выше, чем выше частота мутантного аллеля и степень его вредоносности (s). Очевидно, что скорость появления в популяции мутантных аллелей пропорциональна частоте нормальных, из которых они возникают. Пренебрегая обратными мутациями, т. е. возникновением нормальных аллелей из мутантных, повышение доли аномальных аллелей за одно поколение можно оценить с помощью уравнения Δq = vp, (3.22) где v — мутабильность локуса, р — частота нормального аллеля. В то же время очищающий отбор снижает частоту мутантных аллелей со скоростью 0,5pqs за поколение (где s — средний коэффициент отбора против вредных мутаций). Поскольку оба процесса — мутирование и отбор — противоположны по своему эффекту на приспособленность популяции, то должна существовать точка мутационно-селекционного равновесия, когда поступление новых мутаций компенсируется их удалением из популяции очищающим отбором. Очевидно, в этой точке должно выполняться приближенное равенство vp = 0,5pqs, откуда можно оценить равновесную частоту вредного аллеля q:
 (3.23)
Эта частота прямо пропорциональна скорости появления вредных мутаций и обратно пропорциональна их среднему коэффициенту отбора. Поскольку природные популяции, несмотря на постоянно действующий процесс мутирования, не вымирают, то, очевидно, очищающий отбор со своей задачей успешно справляется. Между прочим, прямая оценка частот вредных мутаций в природных популяциях дрозофилы согласуется с уравнением (3.23).
Дрейф генов
Легко догадаться, что размер любой природной популяции не бесконечен хотя бы из-за ограниченности пищевых ресурсов. Ясно, что колебания в интенсивности таких факторов, как хищники, болезни и т. д., должны отражаться на численности особей. Тем не менее популяция, обитающая в одной и той же местности в течение нескольких тысяч лет, способна сохранять свою численность на более или менее постоянном уровне. Чтобы лучше уловить идею генетического дрейфа, примем, что размер популяции неизменен. Мы знаем, что каждая особь способна произвести (и действительно производит) очень много гамет, которые после оплодотворения могли бы дать начало большому числу потомков. Постоянство размера популяции означает, что в среднем одна особь производит всего две гаметы, дающих начало взрослым особям следующего поколения. Иными словами, в основе формирования генофонда популяции потомков лежит выбор 2N гамет из гигантского по объему гаметного фонда родителей. Насколько полно аллели этих выбранных гамет отражают генофонд популяции?
При случайном выборе из гигантского гаметного фонда всего 2N гамет (пусть даже ровно по две от каждой особи) аллельные частоты в генофонде потомков обязательно отклонятся от их значений в генофонде родителей. Отметим, что при случайном выборе двух гамет из всех производимых одной гетерозиготной особью мы точно отразим ее генотип лишь в 50 % случаев. В остальных 50 % обе гаметы будут нести только один аллель, другой будет потерян.
Интуитивно ясно, что при уменьшении объема гаметной выборки падает точность, с которой она отражает аллельный состав генофонда родителей. Специальный анализ показывает, что наиболее вероятное (среднеквадратическое) отклонение (σ) частоты аллеля в обе стороны от исходной величины растет с уменьшением размера популяции, подчиняясь соотношению
(3.23)
Эта частота прямо пропорциональна скорости появления вредных мутаций и обратно пропорциональна их среднему коэффициенту отбора. Поскольку природные популяции, несмотря на постоянно действующий процесс мутирования, не вымирают, то, очевидно, очищающий отбор со своей задачей успешно справляется. Между прочим, прямая оценка частот вредных мутаций в природных популяциях дрозофилы согласуется с уравнением (3.23).
Дрейф генов
Легко догадаться, что размер любой природной популяции не бесконечен хотя бы из-за ограниченности пищевых ресурсов. Ясно, что колебания в интенсивности таких факторов, как хищники, болезни и т. д., должны отражаться на численности особей. Тем не менее популяция, обитающая в одной и той же местности в течение нескольких тысяч лет, способна сохранять свою численность на более или менее постоянном уровне. Чтобы лучше уловить идею генетического дрейфа, примем, что размер популяции неизменен. Мы знаем, что каждая особь способна произвести (и действительно производит) очень много гамет, которые после оплодотворения могли бы дать начало большому числу потомков. Постоянство размера популяции означает, что в среднем одна особь производит всего две гаметы, дающих начало взрослым особям следующего поколения. Иными словами, в основе формирования генофонда популяции потомков лежит выбор 2N гамет из гигантского по объему гаметного фонда родителей. Насколько полно аллели этих выбранных гамет отражают генофонд популяции?
При случайном выборе из гигантского гаметного фонда всего 2N гамет (пусть даже ровно по две от каждой особи) аллельные частоты в генофонде потомков обязательно отклонятся от их значений в генофонде родителей. Отметим, что при случайном выборе двух гамет из всех производимых одной гетерозиготной особью мы точно отразим ее генотип лишь в 50 % случаев. В остальных 50 % обе гаметы будут нести только один аллель, другой будет потерян.
Интуитивно ясно, что при уменьшении объема гаметной выборки падает точность, с которой она отражает аллельный состав генофонда родителей. Специальный анализ показывает, что наиболее вероятное (среднеквадратическое) отклонение (σ) частоты аллеля в обе стороны от исходной величины растет с уменьшением размера популяции, подчиняясь соотношению
 (3.24)
т. е. это отклонение обратно пропорционально квадратному корню из размера популяции.
Итак, на основании чисто статистических закономерностей частота каждого аллеля при смене поколений претерпевает сдвиг в совершенно непредсказуемом направлении. Этот феномен и представляет собой генный дрейф, его статистический характер не позволяет точно предсказать значения аллельных частот в популяции в ряду поколений. Более того, динамику изменения этих частот невозможно воспроизвести в принципе. Сколько бы раз мы ни следили за частотой какого-то аллеля, мы никогда не встретимся с двумя одинаковыми его судьбами.
Особое значение генный дрейф приобретает в вопросе о судьбе аллелей, не различимых естественным отбором. Такие аллели принято называть селективно-нейтральными или просто нейтральными. Сейчас мы увидим, что дрейф генов выступает как мощный фактор фиксации нейтральных аллелей. Проследим за динамикой частоты такого аллеля (A1).
Пусть в популяции (будем считать ее популяцией нулевого поколения) исходная частота этого аллеля равна р0.
Предположим, у нас есть возможность очень много раз проследить за динамикой изменения частоты аллеля А1 в большом ряду поколений. Тогда, определяя его частоту (р1) в популяциях первого поколения, в каждой из них мы обнаружим отклонение от значения р0. Поэтому частоту аллеля А1 в любой из популяций первого поколения можно представить как р0 + Δр (Δр — сдвиг частоты). Поскольку естественный отбор аллели не различает, вероятность изменения частоты аллеля А1 в обоих направлениях одна и та же, т. е. среднее значение сдвига (где усреднение проведено по всем популяциям первого поколения) должно равняться нулю. Иными словами, средняя частота данного аллеля в совокупности популяций первого поколения <р1> должна в точности равняться его исходной частоте в нулевом поколении.
Так как направление сдвига частоты аллеля А1 сохраняет свою полную неопределенность и в последующих поколениях, то средняя частота этого аллеля <р2> в совокупности популяций второго поколения не должна отличаться от <р1>. Отсюда мы приходим к выводу, что в любом поколении средняя частота аллеля в совокупности популяций, происходящих от какой-то одной, остается равной частоте этого аллеля в исходной популяции, т. е.
р0 = <р1> = <р2> =… (3.25)
Несмотря на то что в каждой популяции динамика изменения аллельных частот непредсказуема, мы можем с полной уверенностью утверждать, что рано или поздно такие изменения прекратятся. Могут быть только два финала: или частота аллеля возрастет до единицы, или она опустится до нуля. Достижение нуля означает потерю аллеля, а достижение единицы — его фиксацию. На рис. 11 приведено несколько судеб нейтральных аллелей, смоделированных компьютером. Каждая траектория частоты аллеля заканчивается либо нулем, либо единицей. Случайный характер изменения аллельной частоты напоминает движение броуновской частицы в трубке, на концы которой помещено клейкое вещество. Очевидно, что при своем блуждании частица когда-нибудь достигнет одного из концов трубки и там приклеится (зафиксируется). Итак, генный дрейф «стремится» уменьшить разнообразие аллелей, одна их часть теряется, а другая фиксируется.
Теперь рассмотрим совокупность всевозможных финалов частоты аллеля А1. У части финальных популяций она будет равна единице, а у всех других — нулю. Очевидно, что доля популяций, зафиксировавших в конце концов аллель А1 (иначе говоря, вероятность фиксации данного аллеля), равна его средней частоте в полной совокупности финальных популяций. Поскольку (см. формулу (3.25)) последняя должна равняться частоте аллеля в исходной популяции (р0), мы приходим к нетривиальному заключению: вероятность фиксации нейтрального аллеля равна его частоте в генофонде популяции. Например, если эта частота равна 0,5, то аллель с равной вероятностью может или исчезнуть из популяции, или в ней зафиксироваться. Теперь перейдем к очень важному следствию — к судьбе только что возникшей мутации.
(3.24)
т. е. это отклонение обратно пропорционально квадратному корню из размера популяции.
Итак, на основании чисто статистических закономерностей частота каждого аллеля при смене поколений претерпевает сдвиг в совершенно непредсказуемом направлении. Этот феномен и представляет собой генный дрейф, его статистический характер не позволяет точно предсказать значения аллельных частот в популяции в ряду поколений. Более того, динамику изменения этих частот невозможно воспроизвести в принципе. Сколько бы раз мы ни следили за частотой какого-то аллеля, мы никогда не встретимся с двумя одинаковыми его судьбами.
Особое значение генный дрейф приобретает в вопросе о судьбе аллелей, не различимых естественным отбором. Такие аллели принято называть селективно-нейтральными или просто нейтральными. Сейчас мы увидим, что дрейф генов выступает как мощный фактор фиксации нейтральных аллелей. Проследим за динамикой частоты такого аллеля (A1).
Пусть в популяции (будем считать ее популяцией нулевого поколения) исходная частота этого аллеля равна р0.
Предположим, у нас есть возможность очень много раз проследить за динамикой изменения частоты аллеля А1 в большом ряду поколений. Тогда, определяя его частоту (р1) в популяциях первого поколения, в каждой из них мы обнаружим отклонение от значения р0. Поэтому частоту аллеля А1 в любой из популяций первого поколения можно представить как р0 + Δр (Δр — сдвиг частоты). Поскольку естественный отбор аллели не различает, вероятность изменения частоты аллеля А1 в обоих направлениях одна и та же, т. е. среднее значение сдвига (где усреднение проведено по всем популяциям первого поколения) должно равняться нулю. Иными словами, средняя частота данного аллеля в совокупности популяций первого поколения <р1> должна в точности равняться его исходной частоте в нулевом поколении.
Так как направление сдвига частоты аллеля А1 сохраняет свою полную неопределенность и в последующих поколениях, то средняя частота этого аллеля <р2> в совокупности популяций второго поколения не должна отличаться от <р1>. Отсюда мы приходим к выводу, что в любом поколении средняя частота аллеля в совокупности популяций, происходящих от какой-то одной, остается равной частоте этого аллеля в исходной популяции, т. е.
р0 = <р1> = <р2> =… (3.25)
Несмотря на то что в каждой популяции динамика изменения аллельных частот непредсказуема, мы можем с полной уверенностью утверждать, что рано или поздно такие изменения прекратятся. Могут быть только два финала: или частота аллеля возрастет до единицы, или она опустится до нуля. Достижение нуля означает потерю аллеля, а достижение единицы — его фиксацию. На рис. 11 приведено несколько судеб нейтральных аллелей, смоделированных компьютером. Каждая траектория частоты аллеля заканчивается либо нулем, либо единицей. Случайный характер изменения аллельной частоты напоминает движение броуновской частицы в трубке, на концы которой помещено клейкое вещество. Очевидно, что при своем блуждании частица когда-нибудь достигнет одного из концов трубки и там приклеится (зафиксируется). Итак, генный дрейф «стремится» уменьшить разнообразие аллелей, одна их часть теряется, а другая фиксируется.
Теперь рассмотрим совокупность всевозможных финалов частоты аллеля А1. У части финальных популяций она будет равна единице, а у всех других — нулю. Очевидно, что доля популяций, зафиксировавших в конце концов аллель А1 (иначе говоря, вероятность фиксации данного аллеля), равна его средней частоте в полной совокупности финальных популяций. Поскольку (см. формулу (3.25)) последняя должна равняться частоте аллеля в исходной популяции (р0), мы приходим к нетривиальному заключению: вероятность фиксации нейтрального аллеля равна его частоте в генофонде популяции. Например, если эта частота равна 0,5, то аллель с равной вероятностью может или исчезнуть из популяции, или в ней зафиксироваться. Теперь перейдем к очень важному следствию — к судьбе только что возникшей мутации.
 Рис. 11. Дрейф аллельных частот в пяти машинных экспериментах. Размер популяции N = 10. Исходная частота обоих аллелей 0,5 (по: [Kimura, 1983]).
Рис. 11. Дрейф аллельных частот в пяти машинных экспериментах. Размер популяции N = 10. Исходная частота обоих аллелей 0,5 (по: [Kimura, 1983]).
Пусть в популяции возник совершенно новый нейтральный аллель. Очевидно, что его частота в момент возникновения равна 1/2N, поэтому и зафиксироваться с помощью дрейфа он может с той же вероятностью. Этот результат приводит нас к грустному выводу: фактически любой вновь возникший аллель обречен на исчезновение, поскольку вероятность этого события чрезвычайно близка к единице (1 — 1/2N). В то же время заметим, что процесс мутирования постоянно ведет к появлению новых аллелей. Если через V обозначить мутабильность локуса, то в каждом поколении в популяции должно появляться 2NV мутантных аллелей. Поскольку они не различаются естественным отбором, любой из них может зафиксироваться с вероятностью 1/2N. Значит, в данном случае скорость фиксации любого мутантного аллеля только за счет генного дрейфа должна равняться 2NV(1/2N). Обозначив скорость фиксации мутантного аллеля через К, приходим к очень простому уравнению K = V. (3.26) Этот изящный результат, полученный М. Кимурой в 1960 г., показывает, что в популяциях (вне зависимости от их размеров) может идти процесс фиксации новых аллелей, скорость которого определяется только скоростью мутирования локусов. Однако среднее число поколений, нужное для фиксации мутантного аллеля с помощью дрейфа, пропорционально размеру популяции и, как показал Кимура, оно близко к 4N. Заметим, что геном любого вида содержит несколько тысяч локусов, поэтому в генофонде любой популяции только вследствие неизбежного генного дрейфа почти в каждом поколении фиксируются какие-то аллели. Однако подчеркнем, что этот вывод мы получили при допущении, что жизнеспособность организма не зависит от аллельного состава локусов; иными словами, естественный отбор эти аллели не различает. Несмотря на то что фиксация нейтральных аллелей занимает много времени, у данного процесса есть одна выгодная для популяции особенность — он проходит для нее совершенно бескровно. Движущий отбор До сих пор мы имели дело с панмиктической популяцией конечного размера, занимающей ограниченную территорию с фиксированными условиями среды. На самом же деле мир, в котором обитает популяция, изменчив, и идеально приспособиться к нему невозможно. Представим себе, что наша популяция в какой-то момент времени хорошо приспособлена к условиям своего существования, однако вдруг эти условия начали медленно, но неуклонно ухудшаться. В данной ситуации перед популяцией встает проблема огромной сложности: нужно не просто отбраковать относительно редкие мутантные аллели (чему в немалой степени помогает генетический дрейф), но и заменить старые, далеко не дефектные аллели на иные, более благоприятные, т. е. лучше соответствующие новым условиям среды. Будем считать, что благоприятные мутации полудоминантны, т. е. h = 0,5. Если принять приспособленность гомозигот по старому аллелю (w11) за единицу, то приспособленность гомозигот по благоприятному аллелю (w22) должна равняться (1+s), а гетерозигот — (1+0,5s). По аналогии с рассмотренным выше случаем вредных мутаций, заменяя — s на +s, можно рассчитать среднюю приспособленность популяции (w) и среднюю приспособленность доноров благоприятного аллеля (w2): w = 1 + qs, (3.27) w2 = 1 + 0,5ps + qs, (3.28) тогда сдвиг частоты благоприятного аллеля за одну генерацию можно определить по формуле Δq = (0,5pqs) / (1 + qs). (3.29) При малых значениях q и s знаменатель в правой части этого уравнения мало отличается от единицы, поэтому с хорошей точностью должно выполняться соотношение Δq ≈ 0,5pqs, (3.30) т. е. скорость возрастания частоты благоприятного аллеля в генофонде популяции прямо пропорциональна его частоте и селективному преимуществу. Глядя на соотношение (3.30), можно подумать, что были бы в наличии благоприятные мутации, а процесс ихфиксации — дело десятое. Однако анализ решения этого уравнения показывает, что даже при таком высоком коэффициенте отбора, как 0,1, для достижения частот благоприятного аллеля, близких к единице, требуется около 100 поколений, при s = 0,01 — не менее 1000. В этих оценках мы не учитывали ограниченность размера реальных популяций. Кроме того, мы полагали, что каждый носитель благоприятной мутации имеет все возможности реализовать свое селективное преимущество. На самом же деле, разбиение вида на небольшие популяции является весьма эффективным средством не столько для фиксации, сколько для удаления редких аллелей — как нейтральных, так и благоприятных. Для того чтобы новый благоприятный аллель вытеснил старый, нужно еще и простое везение. К сожалению, мы сейчас убедимся, что шансы на такое везение не очень велики. Представим себе, что в популяции сколь угодно большого, но постоянного размера возник благоприятный аллель А2. Его носителем может быть только гетерозигота А1А2. Поскольку размер популяции постоянный, каждая особь передает в генофонд следующего поколения в среднем только две копии локуса А. Значит, гетерозигота А1А2 может передать эти две копии в виде одной из трех комбинаций: А1А1, А2А2 и А1А2. Первая комбинация означает потерю аллеля А2, вероятность этого события равна 25 %. В 50 % случаев в генофонд следующего поколения будет передана только одна копия мутантного аллеля (и в следующем поколении он может снова с вероятностью 25 % потеряться), а в 25 % случаев количество мутантных аллелей удвоится. В этом рассуждении мы не учитывали эффекта статистического варьирования числа потомков каждого родителя. Точное решение задачи было получено выдающимся английским математиком и генетиком Р. Фишером. Он продемонстрировал, что вероятность потери вновь возникшей нейтральной мутации в первом же поколении равна 0,37 и быстро растет с увеличением числа поколений, стремясь в конечном счете к единице. Если же мутантный аллель сообщает его носителю селективное преимущество s в гомозиготном состоянии и 0,5s в гетерозиготном, то вероятность его фиксации численно близка к s. Итак, если коэффициент отбора в пользу носителя одной копии благоприятного аллеля составляет целых 5 %, то все-таки шансы его утратить равны 90 %. Следует заметить, что обычно s намного меньше единицы и даже меньше 0,1. Положение может спасти лишь повторное возникновение благоприятных мутаций. Если скорость их возникновения равна v, то в популяции с размером N за одно поколение должно появляться 2Nv новых благоприятных мутаций, из которых будет фиксироваться доля, численно равная s. Отсюда скорость фиксации полудоминантных благоприятных мутаций (K) определяется уравнением К = 2Nvs. (3.31) Заметим, что здесь v обозначает не скорость мутирования вообще, а только скорость возникновения благоприятных мутаций. Если среди всех возникающих за поколение мутаций (V) благоприятные составляют долю f, то уравнение (3.31) можно слегка преобразовать: К = 2NVfs. (3.32) Эта формула хорошо передает особенность адаптивной эволюции. Ведь если число генов в геномах разных видов различается незначительно, то при приблизительном равенстве мутабильности локусов скорость адаптивной эволюции прямо пропорциональна доле благоприятных мутаций (среди всех возникающих) и средней степени их благоприятности. Если макроэволюция полностью сводима к микроэволюции, то уравнение (3.32) показывает, что для ускорения эволюции есть только два средства: увеличить селективное преимущество новых аллелей над старыми (s) и, главное, повысить «выход» благоприятных мутаций (f). Плата за эволюцию До сих пор мы рассматривали приспособленность как шансы зигот стать взрослыми организмами. Однако это понятие можно трактовать чуть-чуть иначе. В популяции постоянных размеров каждая особь оставляет в среднем одного взрослого потомка. Если особи с каким-то генотипом имеют повышенную или пониженную приспособленность, то это значит, что среднее число их потомков должно быть несколько больше или меньше единицы. Получается, что в популяции фиксированных размеров число взрослых потомков, оставляемых особью (ее мальтузианский параметр), может служить мерой ее приспособленности. Мы знаем, что приспособленность популяции, в генофонде которой присутствует благоприятный аллель, за одно поколение немного возрастает, следуя за увеличением частоты данного аллеля. Но так как биомасса популяции увеличиться не может, то для сохранения прежней численности (в среднем один потомок на одного родителя) должно сократиться число потомков от обычных особей, не обладающих благоприятным аллелем. Иными словами, какая-то часть популяции должна остаться без взрослых потомков, что с генетической точки зрения эквивалентно ее гибели (случай так называемой генетической смерти). Очевидно, что это падение относительной приспособленности должно компенсироваться увеличением мальтузианского параметра у носителей благоприятного аллеля. Процесс фиксации благоприятного аллеля обычно занимает много поколений, и на каждом из них имеет место гибель определенной доли популяции. Просуммировав эти доли по всем поколениям (с момента появления аллеля до его фиксации), можно получить величину так называемой платы за эволюцию. Количественную оценку этой суммы дал в 1957 г. знаменитый английский ученый-энциклопедист Дж. Холдейн. Он продемонстрировал, что плата за фиксацию полудоминантного благоприятного аллеля равна -2lnq0 (q0 — начальная частота аллеля). Иными словами, эта плата не зависит от коэффициента отбора, имеет значение лишь исходная частота благоприятного аллеля. Если размер популяции равен миллиону особей, то частота благоприятного аллеля в момент его возникновения (1/2N) должна быть равна 0,5 * 10-6. Удвоенный натуральный логарифм этой величины равен примерно 29, т. е. для реальных популяций его можно принять равным 30. Заметим, что снижение средней приспособленности на единицу означает генетическую смерть (не надо смешивать ее с обычной смертью) всех особей популяции. Получается, что за фиксацию (с помощью отбора) одного аллеля популяция должна «заплатить» 30-кратной генетической гибелью своих особей. Это образное выражение совсем не следует понимать буквально. Биомасса эволюционирующей популяции остается на прежнем уровне, так как снижение относительной приспособленности носителей старого аллеля компенсируется повышением мальтузианского параметра у особей с благоприятным аллелем. Однако такое повышение не может превзойти возможностей особи к размножению — ее репродуктивного потенциала. Это обстоятельство ставит естественное ограничение на скорость адаптивной эволюции. Необходимость постоянно компенсировать генетическую гибель части популяции означает, образно выражаясь, увеличение нагрузки на репродуктивную функцию особей (во всяком случае, носителей благоприятного аллеля). Теперь представим, что в популяции идет процесс фиксации сразу нескольких благоприятных аллелей. Ясно, что в этом случае плата за адаптивную эволюцию резко возрастает, что заставляет нас прийти к весьма пессимистическому выводу: популяция не в состоянии осуществлять параллельные адаптивные замещения аллелей сразу по нескольким локусам, поскольку дополнительная нагрузка на репродуктивный потенциал может превзойти возможности организма. Выходит, что благоприятные мутации скорее всего фиксируются популяцией не параллельно, а последовательно, т. е. адаптивная эволюция не может идти быстро. Определенным выходом из «дилеммы Холдейна» могло бы явиться кооперативное, синергическое взаимодействие благоприятных мутаций. Подобное взаимодействие подразумевает, что две такие мутации повышают приспособленность больше, чем на простую сумму соответствующих коэффициентов отбора. В этом случае естественный отбор «хорошо видит» лишь комбинацию обоих мутантных генов и поэтому фиксирует их единым пакетом. Генетический груз В любой природной популяции всегда присутствуют вредные аллели, снижающие уровень ее приспособленности. Очищающий отбор «стремится» эти аллели удалить, но они снова появляются за счет вечно идущего процесса мутирования. Выше мы рассмотрели условия существования мутационно-селекционного равновесия. Для полудоминантных мутаций их равновесная частота довольно низка. Если в каком-то локусе скорость возникновения вредных мутаций равна 10-6, а s ~ 0,03, то равновесная частота вредящих аллелей в генофонде составит всего 6*10-5 (см.(3.23)). Казалось бы ничтожная доля, но ведь наше рассуждение подходит для всех локусов генома. Если это число равно 10 тысячам, то равновесная частота поврежденных аллелей при суммировании по всем локусам будет близка к 2/3. А если средняя скорость мутирования выше? Например, если средняя мутабильность локуса равна 10-5, то можно прийти к заключению, что в природных популяциях вообще отсутствуют особи без дефектных аллелей. Исследования японского генетика Т. Мукаи, проведенные на дрозофиле, свидетельствуют в пользу именно этого предположения. Мукаи проанализировал спектр мутаций, снижающих жизнеспособность мух, и обнаружил, что большая их часть представлена полудоминантными мутациями, снижающими приспособленность всего на 2–4 %, остальные мутации рецессивны и снижают в гомозиготном состоянии жизнеспособность мух практически до нуля. Общая скорость возникновения слегка вредящих мутаций составляет около 0,4 на гаплоидный геном (т. е. на гамету) за поколение. Заметим, что Мукаи оценивал лишь мутации, снижающие жизнеспособность, тогда как приспособленность должна включать и другие ее компоненты (например, плодовитость). В связи с этим реальное число повреждающих мутаций, возникающих в каждом поколении в геноме гаметы, может оказаться близким к единице (если не больше). При такой интенсивности мутирования можно уже подумать об уровне приспособленности популяции. Если мутантов не слишком много, то понижение производства ими потомков вполне может быть скомпенсировано усиленным размножением особей, лишенных генетических недостатков. Этих последних назовем для удобства элитой. Различие между числом потомков у элиты и среднестатистической особи отражает так называемый мутационный генетический груз (L), смысл которого ясен из формулы L = (wэ — w) / wэ, (3.33) где wэ — средняя приспособленность генетической элиты. Фактически генетический груз показывает, во сколько раз особь из элиты превосходит по мальтузианскому параметру среднюю особь популяции. Теперь снова вернемся к реальной ситуации, когда в среднем каждая гамета получает одну вредную мутацию. Как с этим может справиться очищающий отбор? Здесь может показаться, что положение совершенно безвыходное, поскольку все гаметы оказываются бракованными, и элите не из чего возникнуть. Однако дело спасает случайный характер мутирования. Распределение мутаций по гаметам равносильно распределению капелек дождя по плиткам мостовой. Пусть дождь был слабым, и на одну плитку в среднем упала одна капля, но это не означает, что на каждую плитку попало в точности по одной капле. Ясно, что на некоторые плитки упали две, на иные три капли, а часть плиток вообще осталась сухой. В теории вероятностей такая ситуация хорошо описывается законом Пуассона. В соответствии с ним доля сухих плиток равна е-u. Здесь е — основание натуральных логарифмов, и — среднее число капель дождя, приходящееся на одну плитку. В нашем генетическом случае и — это число вредных мутаций, возникающее в среднем в одной гамете, поэтому доля гамет, счастливо избежавших мутаций, равняется е-u, а доля зигот, возникших от слияния бездефектных гамет, составляет е-2и. Отсюда следует, что для сохранения средней приспособленности популяции бездефектные особи должны размножиться в е2и раз. В частности, при и = 1 каждая особь должна произвести не менее 7,39 потомков, что составляет довольно приличную нагрузку на репродуктивную способность элитных самок. А если эта величина еще больше?… Однако вредные мутации могли бы удаляться не штуками, а сразу пакетами — по две, по три и т. д. Например, если вредная мутация рецессивна, то в гетерозиготном состоянии она не сказывается на приспособленности. Поэтому отбор «видит» только мутантную гомозиготу и, удаляя ее, освобождает генофонд сразу от двух экземпляров вредного аллеля. Если вредящий эффект мутации ощущается уже у гетерозиготы, то ее гибель унесет один вредный аллель, но заодно и один хороший. Следовательно, учитывая весьма широкое распространение слегка вредящих полудоминантных мутаций, природные популяции должны быть обременены весьма существенным генетическим грузом. Средством для его снижения мог бы быть кооперативный характер взаимодействия вредных аллелей разных локусов, когда вредящий эффект одной и той же мутации тем выше, чем больше подобных мутаций уже присутствует в геноме. При этом приспособленность особи по мере добавления мутаций падает по очень крутой параболе. Получается, что геном особи как бы накапливает вредные мутации, а при достижении какого-то порогового значения происходит резкое падение приспособленности; особь с таким геномом гибнет, освобождая генофонд популяции сразу от нескольких вредных мутаций. По мысли советского генетика А.С. Кондрашова, деление особей на два пола является специальным изобретением для снижения генетического груза. Его расчеты показывают, что половой процесс, резко увеличивая размах изменчивости особей по числу вредных мутаций, позволяет популяции гораздо эффективнее освобождаться от последних. Главный вывод, который мы должны сделать после ознакомления с проблемой генетического груза, заключается в том, что среднее число вредных мутаций, возникающих в одной гамете за поколение, не должно превышать определенной фиксированной величины и* (которая является произведением средней мутабильности генов на их число в геноме). Скорее всего уровень спонтанного мутирования в природных популяциях близок к этой критической величине. В пользу такого предположения свидетельствуют две группы фактов. Во-первых, скорость появления однотипных мутаций с качественным эффектом на фенотип у совершенно разных организмов довольно близка; во-вторых, оценка числа жизненно важных генов (потеря которых ведет к прекращению развития) в геноме таких эволюционно далеких видов, как дрозофила и мышь, дает в обоих случаях близкие значения (5—10 тысяч генов). Заметим, что наличие верхнего предела в скорости мутирования (u*) равносильно существованию ограничения на рост информационного содержания генома. Это обстоятельство часто не учитывается многими молекулярными биологами, которые считают, что количество наследственной информации можно оценить исходя из содержания ДНК в гаплоидном ядре. Медленные сдвиги среды Итак, судьба разобщенных барьерами частей вида определяется соотношением двух процессов — экологической дифференциацией популяций и разрушением изолирующих барьеров. Находясь в несколько отличных экологических условиях, популяции под влиянием естественного отбора «стремятся» к ним лучше приспособиться, т. е. по ряду локусов зафиксировать разные аллели. Но этому стремлению противодействует генный поток, который, «просачиваясь» сквозь несовершенные барьеры, может вносить в каждую популяцию неблагоприятные (т. е. менее отвечающие местным условиям) аллели. Таким образом, генный поток выступает здесь как фактор, понижающий приспособленность каждой популяции, что позволяет говорить о миграционном генетическом грузе. Есть только одно средство для закрепления успеха экологической дифференциации разделенных популяций — они должны за время своей изоляции стать разными видами, т. е. заменить географический барьер генетическим. Постоянное «стремление» вида расщепиться на несколько дочерних, по существу, представляет собой попытку вида освободиться от миграционного генетического груза. Кроме того, на периферии своего ареала вид может сталкиваться с качественно иными условиями среды, адаптация к которым означала бы прорыв вида в новую экологическую область. Обрисованная здесь картина выглядит довольно оптимистично, но не следует забывать, что все эти адаптационные процессы предполагает фиксацию новых аллелей по десяткам локусов. Выше мы убедились в том, что на это требуются тысячи и даже десятки тысяч поколений. Но самое главное состоит в том, что естественный отбор проходит без селекционера, а где гарантии, что коэффициент отбора способен сохранять на протяжении нескольких тысяч поколений хотя бы свой знак? Иными словами, для адаптивной эволюции вектор естественного отбора должен сохранять направление и интенсивность в течение огромных отрезков времени. А учитывая противодействие со стороны генного потока, действие естественного отбора должно распространяться на значительную часть ареала вида, желательно на весь его ареал. Естественный отбор — это метафора, его давление можно понимать как постепенное изменение интенсивности каких-то жизненно важных факторов в пространстве и (или) во времени. Например, если ареал вида соседствует с экологической областью, где уровень его абсолютной приспособленности на порядок ниже, то необходимо, чтобы такой контакт сохранялся в течение нескольких десятков тысяч поколений. Или же по всей территории, занятой видом, должно идти однонаправленное изменение какого-то жизненно важного параметра. Для адаптации к такой ситуации необходимо, чтобы скорость изменения параметра была очень небольшой. Во всяком случае, время перехода от оптимальных условий к очень плохим должно составлять тысячи или даже десятки тысяч лет. О значении стабильности среды для экологической дифференциации, по-видимому, свидетельствует удивительное явление повышения численности видов большинства групп растений и животных в направлении от полюсов к экватору. Действительно, климат в тропиках гораздо стабильнее, чем в умеренных широтах. Достаточно вспомнить о недавнем оледенении, практически не отразившемся на животном и растительном мире экваториальной зоны. Кроме того, многие жизненно важные параметры среды испытывают медленные колебания с периодом, исчисляемым миллионами и даже десятками миллионов лет. Так, за последние 220 млн лет климат на Земле сменялся циклически с теплого на холодный не менее 16 раз, примерно по 15 млн лет на цикл (рис. 12). Уровень мирового океана также подвержен циклическим изменениям. Только за последние 65 млн лет его уровень 8 раз изменялся на 100–250 метров. Еще более медленны, но более грандиозны орогенические (горообразовательные) циклы. В них можно выделить длительный геосинклинальный период с продолжительностью до 500 млн лет; за ним в течение примерно 50 млн лет идет период бурного поднятия коры, после которого в течение нескольких десятков миллионов лет может длиться возвращение к геосинклинальному периоду. Во время периода горообразования горы «вздымаются» со скоростью около одного миллиметра в год. В геосинклинальный период эрозия стачивает поднятия быстрее, чем они появляются. Осадки могут накапливаться в мелководных эпиконтинентальных морях со скоростью до одного километра за 10 миллионов лет. Все эти геологические явления, среди которых следует упомянуть еще и дрейф континентов, изменяют, характер морских и воздушных течений, а тем самым и климат многих областей. Итак, данные геологии свидетельствуют о том, что на Земле действительно происходили и происходят очень медленные и однонаправленные изменения среды, захватывающие огромные территории. Характерное время таких изменений выражается миллионами лет.
 Рис. 12. Изменение среднегодовой температуры в течение юры и мела (по: [Красилов, 1977]).
Рис. 12. Изменение среднегодовой температуры в течение юры и мела (по: [Красилов, 1977]).
Легко представить, что чем меньше ареал, занимаемый видом, тем больше у него шансов попасть под действие рассматриваемых здесь медленных однонаправленных сдвигов среды. Американский палеонтолог А. Буко проследил за изменением в течение силура и девона видового состава брахиопод, обитавших в разных геологических провинциях, по-видимому, просто в разных эпиконтинентальных морях. Оказалось, что единственным фактором, от которого зависела скорость изменения фауны, была площадь этих морей. Чем меньше площадь, тем быстрее изменялся состав брахиопод. Заметим, что речь идет о миллиардах особей, занимавших ареалы в миллионы квадратных километров. Рассмотрев в этой главе обстоятельства, при которых возможны ненейтральные изменения генофонда вида, мы приходим к двум важным выводам: во-первых, уровень приспособленности вида к среде следует признать всего лишь удовлетворительным, и, во-вторых, скорость фенотипического сдвига (приспособительного характера) не может быть очень высокой.
Глава 4. Ответ на отбор
Лорд Сомервиль, упоминая о том, чего животноводы достигли по отношению к овце, говорит: «Кажется, будто они начертили на стене форму, совершенную во всех отношениях, и затем придали ей жизнь».Обычно после прочтения книги Ч. Дарвина «Происхождение видов…» и даже после ознакомления с трудами крупнейших неодарвинистов от А. Вейсмана до Э. Майра у читателя остается впечатление, что отбор может все. Ограничения на его творческие возможности накладывают лишь разного рода экологические перипетии, но будь обстоятельства благоприятны, отбор за считанные миллионы лет мог бы в буквальном смысле создать из мухи слона. Со времен Дарвина в качестве главного аргумента в пользу безграничных возможностей отбора выдвигаются успехи человека в создании хозяйственно ценных форм растений и животных. Особенно впечатляют результаты методической селекции последних двух столетий. Чаще всего такая селекция была направлена на повышение мощности вполне конкретной рабочей структуры. Чем не модель для изучения прогрессивной эволюции? Заметим, что скрещивая более и менее продвинутые на пути такого «прогресса» формы и следя за их потомством, можно было бы вскрыть генетическую подоплеку мощности рабочих структур. Итак, если мы действительно хотим проверить тезис о всесильности отбора и научиться измерять его творческие возможности, нам следует совершить небольшой экскурс в генетику количественных признаков. Изменчивость особей Вооружившись подходящим измерительным инструментом, мы можем убедиться, что взрослые особи любой природной популяции отличаются друг от друга по множеству характеристик (признаков): размерам тела и органов, частоте пульса, скорости бега, численности потомства и т. д. Попробуем найти способ, как оценить степень изменчивости популяции по таким мерным признакам. Выберем наугад большое число особей (желательно одного возраста и пола) и измерим у них какую-нибудь характеристику. Ясно, что в любой выборке можно найти одну особь с наименьшей величиной признака и одну — с наибольшей. Разность между этими значениями — диапазон варьирования мерного признака — может служить грубой мерой его изменчивости в популяции. Разобьем диапазон варьирования на ряд равных по величине интервалов. Очевидно, что с помощью этой процедуры мы разделим свою выборку на классы, каждый из которых формируется особями со значением признака, не выходящим за пределы соответствующего интервала. Теперь всю эту совокупность особей можно охарактеризовать ее распределением по признаку, т. е. числом особей, входящих в каждый класс. Фактически мы задаем тем самым математическую функцию численности особей от величины измеряемого параметра.Ч. Дарвин
 Рис. 13. Распределение студентов Гарвардского университета по росту (no: [Castle, 1916]).
Рис. 13. Распределение студентов Гарвардского университета по росту (no: [Castle, 1916]).
Эту функцию удобно воспринимать графически (рис. 13). На оси абсцисс откладываем классовые интервалы, а ордината отражает численность особей, «приписанных» к соответствующему интервалу. К настоящему времени изучены распределения по всевозможным признакам у большого числа популяций разных видов. Оказалось, что их графики обычно представляют собой непрерывные (гладкие) кривые с одной вершиной. Признаки, имеющие такие гладкие одновершинные распределения, в генетике принято называть количественными. Нередко графики этих распределений имеют симметричную колоколообразную форму, напоминая в этом отношении нормальное, или гауссовское, распределение, давно известное в теории вероятностей. Центральным объектом теории вероятностей является так называемая случайная величина, которая характеризуется своим распределением, т. е. вероятностью принимать значения в заданных интервалах. В этой области математики доказывается, что нормальное распределение возникает, когда случайная величина является суммой большого числа независимо варьирующих, но близких по величине слагаемых. Классический пример представляет собой распределение ошибок, изученное великим немецким математиком К. Гауссом. Вся информация о конкретном нормальном распределении заключена в двух его параметрах — среднем значении и дисперсии. Их можно легко вычислить и для нашей выборки особей. Среднее значение <x> — это просто среднеарифметическая величина признака. Угловые скобки обозначают операцию усреднения. Итак,
 (4.1)
где х1, х2…, хn — величина признака у первой, второй и т. д. особей; n — число особей в выборке.
Дисперсия распределения σ2 отражает изменчивость особей по величине признака и является, по определению, средним квадратом отклонения величины признака от его среднего значения:
(4.1)
где х1, х2…, хn — величина признака у первой, второй и т. д. особей; n — число особей в выборке.
Дисперсия распределения σ2 отражает изменчивость особей по величине признака и является, по определению, средним квадратом отклонения величины признака от его среднего значения:
 (4.2)
При получении распределения особей по признаку мы разбиваем всю выборку на n классов, каждый из которых может быть охарактеризован численностью и средним значением признака. Для удобства сравнения разных распределений вместо численности класса лучше взять долю, которую составляют его особи в выборке. Таким образом, i-й класс можно охарактеризовать его долей (рi) в выборке и средним значением признака (хi). В этом случае среднее значение и дисперсия вычисляются по формулам
(4.2)
При получении распределения особей по признаку мы разбиваем всю выборку на n классов, каждый из которых может быть охарактеризован численностью и средним значением признака. Для удобства сравнения разных распределений вместо численности класса лучше взять долю, которую составляют его особи в выборке. Таким образом, i-й класс можно охарактеризовать его долей (рi) в выборке и средним значением признака (хi). В этом случае среднее значение и дисперсия вычисляются по формулам
 (4.3)
(4.3)
 (4.4)
Компоненты дисперсии
Теперь попробуем разобраться в причинах изменчивости особей в природных популяциях. Практический опыт подсказывает, что это явление зависит от среды и от наследственности. То, что среда (в частности, качество питания) влияет на величину количественных признаков, знают все. Влияние наследственности также хорошо известно, достаточно напомнить поговорку: «Яблоко от яблони недалеко падает».
Действие на признак обоих факторов (среды и наследственности), по существу, независимо, что весьма упрощает проблему разложения наблюдаемой (фенотипической) изменчивости на компоненты, связанные с действием каждого фактора. В теории вероятностей доказывается, что дисперсия суммы независимо варьирующих случайных величин является суммой дисперсий каждой из них. Соответственно и фенотипическую дисперсию по величине любого признака (σ2) можно разложить на два слагаемых, на две компоненты — средовую (σе2) и генотипическую (σg2) дисперсии:
(4.4)
Компоненты дисперсии
Теперь попробуем разобраться в причинах изменчивости особей в природных популяциях. Практический опыт подсказывает, что это явление зависит от среды и от наследственности. То, что среда (в частности, качество питания) влияет на величину количественных признаков, знают все. Влияние наследственности также хорошо известно, достаточно напомнить поговорку: «Яблоко от яблони недалеко падает».
Действие на признак обоих факторов (среды и наследственности), по существу, независимо, что весьма упрощает проблему разложения наблюдаемой (фенотипической) изменчивости на компоненты, связанные с действием каждого фактора. В теории вероятностей доказывается, что дисперсия суммы независимо варьирующих случайных величин является суммой дисперсий каждой из них. Соответственно и фенотипическую дисперсию по величине любого признака (σ2) можно разложить на два слагаемых, на две компоненты — средовую (σе2) и генотипическую (σg2) дисперсии:
 (4.5)
В некоторых случаях фенотипическая дисперсия популяции равна средовой. Это происходит тогда, когда генотипы всех особей идентичны. Таковы популяции многих растений, размножающихся бесполым путем — корневищами, побегами и т. д. Среди животных (некоторых рачков, коловраток и даже ящериц) известны клоны партеногенетических самок — потомков одной особи основательницы. Наконец, генетики и селекционеры любят работать с так называемыми изогенными (чистыми) линиями. Особи этих линий (если пренебречь редкими мутациями) генетически идентичны. Тем не менее, как бы мы ни старались выровнять условия среды, нам не удастся добиться полной идентичности их фенотипа.
Дело в том, что онтогенез включает в себя массу многостадийных взаимодействующих между собой процессов, при этом каждая последующая стадия возникает как следствие вполне конкретной предыдущей. Несмотря на хорошую защищенность онтогенеза от внешних воздействий, факторы среды в какой-то мере способны воздействовать на течение любого частного процесса, что может сказаться на прохождении всех последующих стадий и привести к изменению величины любого из многих количественных признаков. Кроме того, протекание многих стадий определяется регуляторными молекулами, число которых относительно невелико, а поэтому подвержено флуктуациям статистического характера. Нельзя же допустить, чтобы все регуляторы были, в свою очередь, регулируемы. Следовательно, сама сложность онтогенеза предопределяет далеко не абсолютную воспроизводимость фенотипа взрослых особей. Традиционно фенотипическую изменчивость генетически идентичных особей называют средовой.
Итак, эффект среды на признак можно рассматривать как сумму большого числа легких толчков, направляющих его развитие с равной вероятностью как в сторону увеличения, так и в сторону уменьшения. В итоге в популяции формируется распределение по признаку, близкое к нормальному. Эти толчки не сдвигают среднего значения признака, которое олицетворяет собой «цель» развития признака, его генотипическое значение. Отсюда вытекает, что среднее значение количественного признака в группе изогенных особей можно рассматривать как его генотипическое значение, которое каждый организм как бы пытается реализовать в ходе своего развития. Тогда как величина средовой дисперсии олицетворяет собой уровень «шума среды», «мешающего» особи реализовать «цель» ее генотипа. Теперь попробуем оценить вклад в фенотипическую изменчивость наследственной компоненты.
Гибридологический анализ количественных признаков
Допустим, мы имеем дело с двумя изогенными линиями Р1 и Р2, принадлежащими одному виду. Несмотря на одинаковые условия среды, их средние значения могут сильно различаться, особенно если линии выделены из географически удаленных популяций. Мы исходим из того, что число генов в геномах сравниваемых линий одинаково, поэтому наследственные различия между ними вызваны разным набором аллелей одних и тех же локусов. Представим себе чисто условно, что в генотипе линии Р1 (с маленьким значением признака) собраны «слабые» аллели, которые будем обозначать строчными буквами, а в генотипе линии Р2 (с большим значением признака) — «сильные» аллели, для их обозначения будем применять прописные буквы.
Пусть разница средних значений признака двух изогенных линий обусловлена отличием в силе аллелей только одного локуса. Тогда генотип линии Р1 обозначим аа, а генотип линии Р2 — АА. Примем еще одно упрощающее условие: пусть по степени доминирования оба аллеля (а и А) равны (h = 0,5). Теперь введем представление об эффекте аллельного замещения. Будем считать, что замещение одного слабого аллеля на сильный увеличивает генотипическое значение признака на α единиц. Тогда замещение обоих слабых аллелей на сильные увеличит это значение на 2α единиц. Вспомним, что генотипическое значение признака равно его среднему значению у особей с одинаковым генотипом, т. е.
(4.5)
В некоторых случаях фенотипическая дисперсия популяции равна средовой. Это происходит тогда, когда генотипы всех особей идентичны. Таковы популяции многих растений, размножающихся бесполым путем — корневищами, побегами и т. д. Среди животных (некоторых рачков, коловраток и даже ящериц) известны клоны партеногенетических самок — потомков одной особи основательницы. Наконец, генетики и селекционеры любят работать с так называемыми изогенными (чистыми) линиями. Особи этих линий (если пренебречь редкими мутациями) генетически идентичны. Тем не менее, как бы мы ни старались выровнять условия среды, нам не удастся добиться полной идентичности их фенотипа.
Дело в том, что онтогенез включает в себя массу многостадийных взаимодействующих между собой процессов, при этом каждая последующая стадия возникает как следствие вполне конкретной предыдущей. Несмотря на хорошую защищенность онтогенеза от внешних воздействий, факторы среды в какой-то мере способны воздействовать на течение любого частного процесса, что может сказаться на прохождении всех последующих стадий и привести к изменению величины любого из многих количественных признаков. Кроме того, протекание многих стадий определяется регуляторными молекулами, число которых относительно невелико, а поэтому подвержено флуктуациям статистического характера. Нельзя же допустить, чтобы все регуляторы были, в свою очередь, регулируемы. Следовательно, сама сложность онтогенеза предопределяет далеко не абсолютную воспроизводимость фенотипа взрослых особей. Традиционно фенотипическую изменчивость генетически идентичных особей называют средовой.
Итак, эффект среды на признак можно рассматривать как сумму большого числа легких толчков, направляющих его развитие с равной вероятностью как в сторону увеличения, так и в сторону уменьшения. В итоге в популяции формируется распределение по признаку, близкое к нормальному. Эти толчки не сдвигают среднего значения признака, которое олицетворяет собой «цель» развития признака, его генотипическое значение. Отсюда вытекает, что среднее значение количественного признака в группе изогенных особей можно рассматривать как его генотипическое значение, которое каждый организм как бы пытается реализовать в ходе своего развития. Тогда как величина средовой дисперсии олицетворяет собой уровень «шума среды», «мешающего» особи реализовать «цель» ее генотипа. Теперь попробуем оценить вклад в фенотипическую изменчивость наследственной компоненты.
Гибридологический анализ количественных признаков
Допустим, мы имеем дело с двумя изогенными линиями Р1 и Р2, принадлежащими одному виду. Несмотря на одинаковые условия среды, их средние значения могут сильно различаться, особенно если линии выделены из географически удаленных популяций. Мы исходим из того, что число генов в геномах сравниваемых линий одинаково, поэтому наследственные различия между ними вызваны разным набором аллелей одних и тех же локусов. Представим себе чисто условно, что в генотипе линии Р1 (с маленьким значением признака) собраны «слабые» аллели, которые будем обозначать строчными буквами, а в генотипе линии Р2 (с большим значением признака) — «сильные» аллели, для их обозначения будем применять прописные буквы.
Пусть разница средних значений признака двух изогенных линий обусловлена отличием в силе аллелей только одного локуса. Тогда генотип линии Р1 обозначим аа, а генотип линии Р2 — АА. Примем еще одно упрощающее условие: пусть по степени доминирования оба аллеля (а и А) равны (h = 0,5). Теперь введем представление об эффекте аллельного замещения. Будем считать, что замещение одного слабого аллеля на сильный увеличивает генотипическое значение признака на α единиц. Тогда замещение обоих слабых аллелей на сильные увеличит это значение на 2α единиц. Вспомним, что генотипическое значение признака равно его среднему значению у особей с одинаковым генотипом, т. е.
 (4.6)
символ < > означает среднее значение признака в линии.
Проведя массовые скрещивания особей таких линий, получим популяцию гибридов F1:
(4.6)
символ < > означает среднее значение признака в линии.
Проведя массовые скрещивания особей таких линий, получим популяцию гибридов F1:
 Все особи популяции F1 имеют один и тот же генотип аА, поэтому изменчивость признака в данном случае обусловлена исключительно средой. Будем считать, что средовая дисперсия для всех популяций (F1, P1 и Р2) одинакова и равна σe2. Среднее значение признака у особей F1 (обозначим его т) должно равняться генотипическому значению гетерозигот аА, т. е. оно должно на α единиц превосходить среднее значение особей линии P1 и настолько же уступать среднему значению особей Р2. Иными словами, m попадает точно в середину интервала между средними значениями признака обеих родительских популяций. Тогда средние значения признака всех рассмотренных популяций можно представить как
<F1> = m; <Р1> = m — а; <Р2> = m + а. (4.7)
Итак, m, m — а и m + а — генотипические значения признака у особей с генетической конституцией аА, аа и АА соответственно.
Перейдем к популяции F2, возникшей или при самооплодотворении, или при панмиктическом скрещивании особей из F1:
Все особи популяции F1 имеют один и тот же генотип аА, поэтому изменчивость признака в данном случае обусловлена исключительно средой. Будем считать, что средовая дисперсия для всех популяций (F1, P1 и Р2) одинакова и равна σe2. Среднее значение признака у особей F1 (обозначим его т) должно равняться генотипическому значению гетерозигот аА, т. е. оно должно на α единиц превосходить среднее значение особей линии P1 и настолько же уступать среднему значению особей Р2. Иными словами, m попадает точно в середину интервала между средними значениями признака обеих родительских популяций. Тогда средние значения признака всех рассмотренных популяций можно представить как
<F1> = m; <Р1> = m — а; <Р2> = m + а. (4.7)
Итак, m, m — а и m + а — генотипические значения признака у особей с генетической конституцией аА, аа и АА соответственно.
Перейдем к популяции F2, возникшей или при самооплодотворении, или при панмиктическом скрещивании особей из F1:
 Из этой схемы видно, что вся совокупность особей F2 разбивается на три генотипических класса; каждый из них можно охарактеризовать его долей в выборке и средним значением признака. Одна четверть особей имеет генотип аа, другая четверть — генотип АА и половина — генотип аА. Поскольку средние значения признака у особей с этими генотипами равны соответственно m — а, m + а и m, то по (4.3) и (4.4) легко рассчитать среднее значение (М) и дисперсию (σg2) для распределения особей по генотипическим классам:
Из этой схемы видно, что вся совокупность особей F2 разбивается на три генотипических класса; каждый из них можно охарактеризовать его долей в выборке и средним значением признака. Одна четверть особей имеет генотип аа, другая четверть — генотип АА и половина — генотип аА. Поскольку средние значения признака у особей с этими генотипами равны соответственно m — а, m + а и m, то по (4.3) и (4.4) легко рассчитать среднее значение (М) и дисперсию (σg2) для распределения особей по генотипическим классам:
 (4.8)
(4.8)
 (4.9)
Таким образом, популяция F2 обладает дисперсией (а2/2), обусловленной различием особей по генотипу. Кроме того, из-за «шума» среды популяция обладает и средовой дисперсией σe2. Этот шум не сдвигает средних значений, поэтому <F2> = m. Эффекты среды и генотипа независимы, отсюда следует, что дисперсия по признаку в поколении F2 должна быть больше средовой на положительную величину а2/2, т. е.
(4.9)
Таким образом, популяция F2 обладает дисперсией (а2/2), обусловленной различием особей по генотипу. Кроме того, из-за «шума» среды популяция обладает и средовой дисперсией σe2. Этот шум не сдвигает средних значений, поэтому <F2> = m. Эффекты среды и генотипа независимы, отсюда следует, что дисперсия по признаку в поколении F2 должна быть больше средовой на положительную величину а2/2, т. е.
 (4.10)
Теперь попробуем рассмотреть более общий случай, когда особи двух изогенных линий различаются аллелями n несцепленных локусов. По-прежнему будем считать, что все слабые аллели собраны у линии P1, а все сильные — у линии Р2. Проведем их скрещивание:
(4.10)
Теперь попробуем рассмотреть более общий случай, когда особи двух изогенных линий различаются аллелями n несцепленных локусов. По-прежнему будем считать, что все слабые аллели собраны у линии P1, а все сильные — у линии Р2. Проведем их скрещивание:
 При оценке среднего значения популяции F1 сделаем два предположения: во-первых, по степени доминирования все аллели равны и, во-вторых, замещение в каждом локусе одного слабого аллеля на сильный увеличивает генотипическое значение признака на одну и ту же величину а. Следовательно, разность средних значений родительских популяций должна быть равна 2na, а среднее значение признака в популяции F, (обозначим его <F1>) будет находиться в точке m, т. е. точно посередине между средними значениями родительских линий. Такая модель, где вклады всех аллелей в величину признака суммируются, получила название аддитивной. Главным основанием для ее применения является попадание среднего значения признака в популяциях F1 и F2 посередине между средними значениями родительских популяций. Итак, для случая n локусов
<F1> = m; <P1> = m — na; <Р2> = m + na. (4.11)
Так как генотип всех особей F1 одинаков, то изменчивость признака в этой популяции обусловлена только влиянием среды, и ее дисперсия равна σe2.
Теперь перейдем к популяции F2, представляющей собой смесь огромного числа (3n) генотипов. Формулу генотипа каждой особи можно записать как ряд из n аллельных пар со случайной комбинацией сильных и слабых аллелей в каждой паре. Поскольку аллельный состав каждого локуса формируется независимо от остальных, то генотипическая дисперсия популяции F2 должна представлять собой сумму дисперсий, каждая из которых отражает варьирование у разных особей числа сильных аллелей в каком-то одном локусе. Напомним, что в данной, аддитивной, модели замещение в любом локусе слабого аллеля на сильный ведет к увеличению генотипического значения признака на одну и ту же величину а. Отсюда следует (см. (4.9)), что каждый из n локусов вносит в генотипическую дисперсию поколения F2 один и тот же вклад а2/2. Итак, величину фенотипической дисперсии σ2 в популяции F2 можно передать формулой
σ2 = σe2 + na2/2. (4.12)
Это равенство вместе с другим
<Р2> — <Р1> = 2na (4.13)
образует систему двух независимых уравнений, позволяющих определить величину п:
При оценке среднего значения популяции F1 сделаем два предположения: во-первых, по степени доминирования все аллели равны и, во-вторых, замещение в каждом локусе одного слабого аллеля на сильный увеличивает генотипическое значение признака на одну и ту же величину а. Следовательно, разность средних значений родительских популяций должна быть равна 2na, а среднее значение признака в популяции F, (обозначим его <F1>) будет находиться в точке m, т. е. точно посередине между средними значениями родительских линий. Такая модель, где вклады всех аллелей в величину признака суммируются, получила название аддитивной. Главным основанием для ее применения является попадание среднего значения признака в популяциях F1 и F2 посередине между средними значениями родительских популяций. Итак, для случая n локусов
<F1> = m; <P1> = m — na; <Р2> = m + na. (4.11)
Так как генотип всех особей F1 одинаков, то изменчивость признака в этой популяции обусловлена только влиянием среды, и ее дисперсия равна σe2.
Теперь перейдем к популяции F2, представляющей собой смесь огромного числа (3n) генотипов. Формулу генотипа каждой особи можно записать как ряд из n аллельных пар со случайной комбинацией сильных и слабых аллелей в каждой паре. Поскольку аллельный состав каждого локуса формируется независимо от остальных, то генотипическая дисперсия популяции F2 должна представлять собой сумму дисперсий, каждая из которых отражает варьирование у разных особей числа сильных аллелей в каком-то одном локусе. Напомним, что в данной, аддитивной, модели замещение в любом локусе слабого аллеля на сильный ведет к увеличению генотипического значения признака на одну и ту же величину а. Отсюда следует (см. (4.9)), что каждый из n локусов вносит в генотипическую дисперсию поколения F2 один и тот же вклад а2/2. Итак, величину фенотипической дисперсии σ2 в популяции F2 можно передать формулой
σ2 = σe2 + na2/2. (4.12)
Это равенство вместе с другим
<Р2> — <Р1> = 2na (4.13)
образует систему двух независимых уравнений, позволяющих определить величину п:
 (4.14)
Хотя эта знаменитая формула Кастла — Райта верна лишь в рамках аддитивной модели, она дает возможность ориентировочно подойти к числу генетических факторов, ответственных за межлинейную разницу величины признака.
Что нам дал этот гибридологический экскурс? Очень много. Хотя природные популяции — это не поколение F2 но и здесь генотипическое значение признака можно считать суммой n независимо варьирующих слагаемых, где n — число локусов в генофонде популяции. Только в отличие от F2 число аллелей каждого локуса в данном случае может быть больше двух, и в пары они соединяются не в отношении 1:2:1, а по закону Харди — Вайнберга. Хотя мы ничего не знаем ни об эффектах этих аллелей, ни о степени их доминирования, ясно одно: популяционная дисперсия признака должна расти с увеличением числа локусов, принимающих участие в его формировании.
Сигма
Очень часто в качестве меры фенотипической изменчивости используют квадратный корень из дисперсии — так называемое среднеквадратичное отклонение (σ). Для экономии места будем именовать эту величину сигмой, по названию греческой буквы, обычно используемой для ее обозначения. Измеряемая в единицах величины самого признака, сигма очень удобна как масштаб для оценки отклонения величины признака от среднепопуляционного значения. Если признак имеет нормальное распределение, то доля особей с отклонением в пределах одной сигмы составляет 68 %, двух сигм — 95 и трех сигм — 99,7 %. В связи с этим полный размах изменчивости признака, распределенного по нормальному закону, попадает в интервал ±3σ (закон трех сигм). В сигмах принято измерять разность средних значений сравниваемых распределений и, в частности, эффект аллельных замещений.
Как и дисперсия, сигма может служить мерой средовой изменчивости — шума среды. Мы видели, что формула Кастла — Райта позволяет приближенно оценить число локусов, ответственных за различие линий по величине количественного признака. Зная это число, можно определить и средний эффект замещений слабого аллеля на сильный. Очень часто эффект оказывается меньше величины средовой сигмы σe. Это обстоятельство позволяет почувствовать основную трудность, с которой сталкивается исследователь генетических основ количественных признаков. Ведь если эффект аллельных замещений меньше шума среды, то по фенотипу особи определить ее генотип невозможно. Это же объясняет и гладкий характер распределений по количественным признакам. Шум среды как бы замазывает генотипические различия особей.
При анализе природных популяций широко используется еще одна мера — коэффициент изменчивости признака, т. е. отношение сигмы к среднему значению, выраженное в процентах. Наиболее поразительная черта этого коэффициента заключается в его стабильности при переходе от популяции к популяции в пределах одного вида и даже при сравнении популяций разных видов одного рода. Более того, коэффициент изменчивости самых разных признаков в популяциях эволюционно далеких видов также не слишком различается, несмотря на большие расхождения в величинах средних значений. Отсюда следует, что между сигмой и средним значением существует связь, близкая к прямой пропорциональности. Однако такой вывод в корне противоречит нашему исходному представлению о нормальном характере распределения особей по признаку.
Напомним, что нормальное распределение однозначно определяется двумя независимыми параметрами — средним значением и дисперсией. Иными словами, если признак распределен в каждой популяции по нормальному закону, то при переходе от одной популяции к другой сигма и среднее значение должны изменяться независимо. Выходит, что наша первоначальная интерпретация средового влияния как суммарного действия большого числа легких независимых толчков не совсем точна. Правда, уже со времен создателя биометрии Ф. Гальтона известно, что при измерении признака логарифмической шкалой связь между сигмой и средним значением зачастую теряется. Если обозначить сдвиг значения признака X, измеренного обычной (арифметической) шкалой, как ΔХ, то отношение этого сдвига к величине признака (его относительный сдвиг) примерно равно изменению признака, измеренного в логарифмическом масштабе, поскольку
ΔX / X ≈ ΔlnX. (4.15)
Распределение, которое становится нормальным после логарифмирования значения случайной величины, называется логарифмически-нормальным. Его характерной особенностью является линейная связь между сигмой и средним значением. Вообще говоря, логарифмически-нормальное распределение совсем не симметрично и обладает длинным «хвостом», плавно спускающимся в сторону увеличения признака. Однако так оно выглядит при большом размахе изменчивости признака в арифметической шкале (например, в 10 раз). Если же такой диапазон значительно скромнее (скажем, только в 1,5 раза), то распределение по признаку в обеих шкалах выглядит очень сходным, и даже в арифметической шкале не слишком отличается от нормального. Поэтому при анализе особей одной популяции, когда размах изменчивости невелик, логарифмически-нормальные распределения выглядят как нормальные.
Опыт биометрии свидетельствует, что наследование количественных признаков, измеренных с помощью логарифмической шкалы, часто удовлетворяет аддитивной модели. Это означает, что межлинейное различие по величине логарифма признака можно трактовать как сумму эффектов аллельных замещений поряду локусов. Заметим, что логарифм числа представим в виде суммы близких по величине слагаемых, когда само это число является произведением близких по значению сомножителей. Выходит, что замещение слабого аллеля на сильный в локусе, ответственном за развитие количественного признака, ведет к увеличению его генотипического значения в какое-то число раз. Причем это число не слишком различается при аллельных замещениях в разных локусах.
Ответ на искусственный отбор
Сначала рассмотрим, что кроется под таким образным понятием, как давление искусственного отбора. Проще всего под ним понимать долю отбракованных особей (I), обычно измеряемую в процентах:
I = (N0 / N)100 %, (4.16)
где N — численность популяции до отбора; N0 — число отбракованных особей. Недостаток формулы (4.16) состоит в отсутствии какой-либо информации о признаке, по которому ведется отбор. Эту сторону работы селекционера передает так называемый селекционный дифференциал (S), который определяется как разница средних значений признака в исходной популяции (до браковки) (М) и у «счастливцев», отобранных для развода (Мс), т. е.
S = Mc — M. (4.16)
Аналогично можно ввести представление об ответе популяции на отбор (R) как о сдвиге среднего значения популяции за одно поколение отбора:
R = M'c — M, (4.18)
где M'c — среднее значение признака у потомков «счастливцев».
Большой экспериментальный материал свидетельствует, что ответ на отбор прямо пропорционален селекционному дифференциалу. Следовательно, чем сильнее давление отбора, тем больше величина ответа на него. Величина этого ответа — внутреннее (генетически обусловленное) свойство популяции. Его количественной мерой является наследуемость — коэффициент пропорциональности (h2), связывающий ответ на отбор с селекционным дифференциалом. Итак, при ответе конкретной популяции на отбор по конкретному количественному признаку выполняется равенство
R = h2S. (4.19)
Наследуемость разных признаков у особей одной популяции может существенно различаться. Кроме того, может различаться и наследуемость одного и того же признака в разных популяциях одного вида. Что же лежит в основе этого явления?
Пусть мы имеем дело с популяцией, состоящей из генетически идентичных особей. Многократно показано, что при любом давлении отбора среднее значение любого признака в следующем поколении практически не изменяется (h2 = 0). Впервые это было продемонстрировано в знаменитом эксперименте В. Иогансена на чистых линиях фасоли. Иными словами, наследуемость равна нулю, если равна нулю генетическая компонента изменчивости.
Представим на момент, что мы в состоянии по фенотипу особи опознать ее генотип. Если к тому же эти особи размножаются бесполым путем, то состав генотипов потомков будет совпадать с таковым у отобранной группы родителей. Следовательно, и средние значения признаков потомков будут совпадать с их средними значениями у родителей, тем самым величина ответа на отбор сравняется с селекционным дифференциалом. Очевидно, что в данном случае h2 = 1.
Обычно же ответ на отбор меньше селекционного дифференциала, так как часть особей попадает в отобранную группу совершенно случайно (вследствие счастливого стечения средовых факторов), поэтому особенность их фенотипа наследоваться не может. Ведь для того, чтобы потомки этих «баловней судьбы» (а не генотипа) попали в отбираемую группу, им снова должно повезти. Но случай на то и случай, чтобы не повторяться регулярно.
В генетике количественных признаков доказывается, что наследуемость равна доле генетической дисперсии в полной фенотипической дисперсии признака. Вспоминая, что эта последняя складывается из генетической и средовой компонент, сказанное можно передать формулой
(4.14)
Хотя эта знаменитая формула Кастла — Райта верна лишь в рамках аддитивной модели, она дает возможность ориентировочно подойти к числу генетических факторов, ответственных за межлинейную разницу величины признака.
Что нам дал этот гибридологический экскурс? Очень много. Хотя природные популяции — это не поколение F2 но и здесь генотипическое значение признака можно считать суммой n независимо варьирующих слагаемых, где n — число локусов в генофонде популяции. Только в отличие от F2 число аллелей каждого локуса в данном случае может быть больше двух, и в пары они соединяются не в отношении 1:2:1, а по закону Харди — Вайнберга. Хотя мы ничего не знаем ни об эффектах этих аллелей, ни о степени их доминирования, ясно одно: популяционная дисперсия признака должна расти с увеличением числа локусов, принимающих участие в его формировании.
Сигма
Очень часто в качестве меры фенотипической изменчивости используют квадратный корень из дисперсии — так называемое среднеквадратичное отклонение (σ). Для экономии места будем именовать эту величину сигмой, по названию греческой буквы, обычно используемой для ее обозначения. Измеряемая в единицах величины самого признака, сигма очень удобна как масштаб для оценки отклонения величины признака от среднепопуляционного значения. Если признак имеет нормальное распределение, то доля особей с отклонением в пределах одной сигмы составляет 68 %, двух сигм — 95 и трех сигм — 99,7 %. В связи с этим полный размах изменчивости признака, распределенного по нормальному закону, попадает в интервал ±3σ (закон трех сигм). В сигмах принято измерять разность средних значений сравниваемых распределений и, в частности, эффект аллельных замещений.
Как и дисперсия, сигма может служить мерой средовой изменчивости — шума среды. Мы видели, что формула Кастла — Райта позволяет приближенно оценить число локусов, ответственных за различие линий по величине количественного признака. Зная это число, можно определить и средний эффект замещений слабого аллеля на сильный. Очень часто эффект оказывается меньше величины средовой сигмы σe. Это обстоятельство позволяет почувствовать основную трудность, с которой сталкивается исследователь генетических основ количественных признаков. Ведь если эффект аллельных замещений меньше шума среды, то по фенотипу особи определить ее генотип невозможно. Это же объясняет и гладкий характер распределений по количественным признакам. Шум среды как бы замазывает генотипические различия особей.
При анализе природных популяций широко используется еще одна мера — коэффициент изменчивости признака, т. е. отношение сигмы к среднему значению, выраженное в процентах. Наиболее поразительная черта этого коэффициента заключается в его стабильности при переходе от популяции к популяции в пределах одного вида и даже при сравнении популяций разных видов одного рода. Более того, коэффициент изменчивости самых разных признаков в популяциях эволюционно далеких видов также не слишком различается, несмотря на большие расхождения в величинах средних значений. Отсюда следует, что между сигмой и средним значением существует связь, близкая к прямой пропорциональности. Однако такой вывод в корне противоречит нашему исходному представлению о нормальном характере распределения особей по признаку.
Напомним, что нормальное распределение однозначно определяется двумя независимыми параметрами — средним значением и дисперсией. Иными словами, если признак распределен в каждой популяции по нормальному закону, то при переходе от одной популяции к другой сигма и среднее значение должны изменяться независимо. Выходит, что наша первоначальная интерпретация средового влияния как суммарного действия большого числа легких независимых толчков не совсем точна. Правда, уже со времен создателя биометрии Ф. Гальтона известно, что при измерении признака логарифмической шкалой связь между сигмой и средним значением зачастую теряется. Если обозначить сдвиг значения признака X, измеренного обычной (арифметической) шкалой, как ΔХ, то отношение этого сдвига к величине признака (его относительный сдвиг) примерно равно изменению признака, измеренного в логарифмическом масштабе, поскольку
ΔX / X ≈ ΔlnX. (4.15)
Распределение, которое становится нормальным после логарифмирования значения случайной величины, называется логарифмически-нормальным. Его характерной особенностью является линейная связь между сигмой и средним значением. Вообще говоря, логарифмически-нормальное распределение совсем не симметрично и обладает длинным «хвостом», плавно спускающимся в сторону увеличения признака. Однако так оно выглядит при большом размахе изменчивости признака в арифметической шкале (например, в 10 раз). Если же такой диапазон значительно скромнее (скажем, только в 1,5 раза), то распределение по признаку в обеих шкалах выглядит очень сходным, и даже в арифметической шкале не слишком отличается от нормального. Поэтому при анализе особей одной популяции, когда размах изменчивости невелик, логарифмически-нормальные распределения выглядят как нормальные.
Опыт биометрии свидетельствует, что наследование количественных признаков, измеренных с помощью логарифмической шкалы, часто удовлетворяет аддитивной модели. Это означает, что межлинейное различие по величине логарифма признака можно трактовать как сумму эффектов аллельных замещений поряду локусов. Заметим, что логарифм числа представим в виде суммы близких по величине слагаемых, когда само это число является произведением близких по значению сомножителей. Выходит, что замещение слабого аллеля на сильный в локусе, ответственном за развитие количественного признака, ведет к увеличению его генотипического значения в какое-то число раз. Причем это число не слишком различается при аллельных замещениях в разных локусах.
Ответ на искусственный отбор
Сначала рассмотрим, что кроется под таким образным понятием, как давление искусственного отбора. Проще всего под ним понимать долю отбракованных особей (I), обычно измеряемую в процентах:
I = (N0 / N)100 %, (4.16)
где N — численность популяции до отбора; N0 — число отбракованных особей. Недостаток формулы (4.16) состоит в отсутствии какой-либо информации о признаке, по которому ведется отбор. Эту сторону работы селекционера передает так называемый селекционный дифференциал (S), который определяется как разница средних значений признака в исходной популяции (до браковки) (М) и у «счастливцев», отобранных для развода (Мс), т. е.
S = Mc — M. (4.16)
Аналогично можно ввести представление об ответе популяции на отбор (R) как о сдвиге среднего значения популяции за одно поколение отбора:
R = M'c — M, (4.18)
где M'c — среднее значение признака у потомков «счастливцев».
Большой экспериментальный материал свидетельствует, что ответ на отбор прямо пропорционален селекционному дифференциалу. Следовательно, чем сильнее давление отбора, тем больше величина ответа на него. Величина этого ответа — внутреннее (генетически обусловленное) свойство популяции. Его количественной мерой является наследуемость — коэффициент пропорциональности (h2), связывающий ответ на отбор с селекционным дифференциалом. Итак, при ответе конкретной популяции на отбор по конкретному количественному признаку выполняется равенство
R = h2S. (4.19)
Наследуемость разных признаков у особей одной популяции может существенно различаться. Кроме того, может различаться и наследуемость одного и того же признака в разных популяциях одного вида. Что же лежит в основе этого явления?
Пусть мы имеем дело с популяцией, состоящей из генетически идентичных особей. Многократно показано, что при любом давлении отбора среднее значение любого признака в следующем поколении практически не изменяется (h2 = 0). Впервые это было продемонстрировано в знаменитом эксперименте В. Иогансена на чистых линиях фасоли. Иными словами, наследуемость равна нулю, если равна нулю генетическая компонента изменчивости.
Представим на момент, что мы в состоянии по фенотипу особи опознать ее генотип. Если к тому же эти особи размножаются бесполым путем, то состав генотипов потомков будет совпадать с таковым у отобранной группы родителей. Следовательно, и средние значения признаков потомков будут совпадать с их средними значениями у родителей, тем самым величина ответа на отбор сравняется с селекционным дифференциалом. Очевидно, что в данном случае h2 = 1.
Обычно же ответ на отбор меньше селекционного дифференциала, так как часть особей попадает в отобранную группу совершенно случайно (вследствие счастливого стечения средовых факторов), поэтому особенность их фенотипа наследоваться не может. Ведь для того, чтобы потомки этих «баловней судьбы» (а не генотипа) попали в отбираемую группу, им снова должно повезти. Но случай на то и случай, чтобы не повторяться регулярно.
В генетике количественных признаков доказывается, что наследуемость равна доле генетической дисперсии в полной фенотипической дисперсии признака. Вспоминая, что эта последняя складывается из генетической и средовой компонент, сказанное можно передать формулой
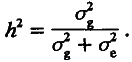 (4.20)
Повторяя процедуру отбора, направленного на изменение величины признака, в течение большого числа поколений мы можем в конце концов добиться фиксации генотипов, включающих в себя только сильные или только слабые аллели (в зависимости от направления отбора). Ясно, что в таком случае весь запас генетической изменчивости будет исчерпан, и дальнейший ответ на отбор станет невозможным.
Теперь обратимся к эксперименту. Действительно, у животных через несколько поколений отбора ответ на него обычно прекращается, и среднее значение признака выходит на плато. Однако интерпретировать это явление не так просто.
Негативные эффекты искусственного отбора
Первый лабораторный эксперимент по отбору на изменение количественных признаков был проведен в начале XX века американским генетиком У. Кастлом. Он ставил перед собой цель — превратить пегих (пятнистых) крыс в одноцветных. В одном варианте отбор был направлен на увеличение площади белых пятен, в другом — в противоположном направлении. Примерно через 20 поколений Кастл был близок к цели, т. е. ему удалось получить почти белых и почти черных крыс, но опыт пришлось прекратить, так как животные захирели и перестали размножаться. Короче говоря, у крыс резко снизились основные показатели приспособленности.
В дальнейшем сходные результаты были получены на большом числе животных объектов — мышах, курах, золотых рыбках, мучных хрущаках и, конечно, на дрозофиле. Везде по мере сдвига самых разных количественных признаков в любом направлении наблюдалось снижение показателей приспособленности: прежде всего жизнеспособности и плодовитости. Популяции как бы сопротивлялись действию искусственного отбора.
Особенно много экспериментов проведено по увеличению числа щетинок на разных частях тела у дрозофилы. Пристрастие именно к этому объекту объясняется очень коротким жизненным циклом плодовой мушки и ее хорошей генетической изученностью. Пример такой селекции приведен на рис. 14. Величина признака сначала монотонно возрастает, затем (обычно после 15–20 поколений отбора) наступает застой, ответ на отбор прекращается, а жизнеспособность и плодовитость мух падают. Оказывается, если отбор прекратить, то основные показатели приспособленности довольно быстро восстанавливаются, однако обычно это сочетается с существенным снижением числа щетинок, т. е. за несколько поколений «отдыха» от отбора величина признака сдвигается назад в сторону его исходных значений. Если после периода «отдыха» отбор возобновить, то можно снова поднять признак на прежний уровень, иногда даже при более высоких значениях главных компонент приспособленности.
Для экспериментов по долговременному отбору на увеличение числа щетинок характерна поразительная невоспроизводимость динамики изменения признака. В то же время в этой картине при всем ее многообразии имеются некоторые общие моменты. Обычно все начинается с довольно быстрого ответа на отбор, потом скорость увеличения признака постепенно падает, и кривая отбора выходит на плато. В этом состоянии «застоя», несмотря на продолжающуюся селекцию, популяция может находиться неопределенно долго. Затем внезапно может наступить фаза быстрого ответа на отбор, и кривая выходит на новое, более высокое плато. Этап нового застоя через какое-то число поколений может опять смениться фазой внезапного ответа, потом снова наступает застой и т. д.
(4.20)
Повторяя процедуру отбора, направленного на изменение величины признака, в течение большого числа поколений мы можем в конце концов добиться фиксации генотипов, включающих в себя только сильные или только слабые аллели (в зависимости от направления отбора). Ясно, что в таком случае весь запас генетической изменчивости будет исчерпан, и дальнейший ответ на отбор станет невозможным.
Теперь обратимся к эксперименту. Действительно, у животных через несколько поколений отбора ответ на него обычно прекращается, и среднее значение признака выходит на плато. Однако интерпретировать это явление не так просто.
Негативные эффекты искусственного отбора
Первый лабораторный эксперимент по отбору на изменение количественных признаков был проведен в начале XX века американским генетиком У. Кастлом. Он ставил перед собой цель — превратить пегих (пятнистых) крыс в одноцветных. В одном варианте отбор был направлен на увеличение площади белых пятен, в другом — в противоположном направлении. Примерно через 20 поколений Кастл был близок к цели, т. е. ему удалось получить почти белых и почти черных крыс, но опыт пришлось прекратить, так как животные захирели и перестали размножаться. Короче говоря, у крыс резко снизились основные показатели приспособленности.
В дальнейшем сходные результаты были получены на большом числе животных объектов — мышах, курах, золотых рыбках, мучных хрущаках и, конечно, на дрозофиле. Везде по мере сдвига самых разных количественных признаков в любом направлении наблюдалось снижение показателей приспособленности: прежде всего жизнеспособности и плодовитости. Популяции как бы сопротивлялись действию искусственного отбора.
Особенно много экспериментов проведено по увеличению числа щетинок на разных частях тела у дрозофилы. Пристрастие именно к этому объекту объясняется очень коротким жизненным циклом плодовой мушки и ее хорошей генетической изученностью. Пример такой селекции приведен на рис. 14. Величина признака сначала монотонно возрастает, затем (обычно после 15–20 поколений отбора) наступает застой, ответ на отбор прекращается, а жизнеспособность и плодовитость мух падают. Оказывается, если отбор прекратить, то основные показатели приспособленности довольно быстро восстанавливаются, однако обычно это сочетается с существенным снижением числа щетинок, т. е. за несколько поколений «отдыха» от отбора величина признака сдвигается назад в сторону его исходных значений. Если после периода «отдыха» отбор возобновить, то можно снова поднять признак на прежний уровень, иногда даже при более высоких значениях главных компонент приспособленности.
Для экспериментов по долговременному отбору на увеличение числа щетинок характерна поразительная невоспроизводимость динамики изменения признака. В то же время в этой картине при всем ее многообразии имеются некоторые общие моменты. Обычно все начинается с довольно быстрого ответа на отбор, потом скорость увеличения признака постепенно падает, и кривая отбора выходит на плато. В этом состоянии «застоя», несмотря на продолжающуюся селекцию, популяция может находиться неопределенно долго. Затем внезапно может наступить фаза быстрого ответа на отбор, и кривая выходит на новое, более высокое плато. Этап нового застоя через какое-то число поколений может опять смениться фазой внезапного ответа, потом снова наступает застой и т. д.
 Рис. 14. Отбор на увеличение числа абдоминальных щетинок у дрозофилы Drosophila melanogaster (по: [Yoo, 1980]).
Рис. 14. Отбор на увеличение числа абдоминальных щетинок у дрозофилы Drosophila melanogaster (по: [Yoo, 1980]).
Вся эта феноменология выглядит весьма загадочно и до сих пор не получила удовлетворительного объяснения. Существует несколько гипотез. Самая простая из них связывает падение приспособленности с так называемой инбредной депрессией. Действительно, обычно численность лабораторных понуляций невелика и в каждом поколении большая их часть подвергается браковке. Поэтому с ходом отбора постепенно повышается степень родства особей, вступающих в скрещивание. А это означает прогрессивное возрастание степени гомозиготности генов, часть которых может быть представлена вредными рецессивными аллелями. Однако расчеты показывают, что инбредная депрессия повинна лишь в части негативных последствий селекции. Вторая гипотеза, высказанная выдающимся американским генетиком и математиком С. Райтом, объясняет негативный эффект селекции нарушением взаимодействия генов, хорошо «подогнанных» друг к другу естественным отбором. В ходе искусственного отбора в генофонде популяции нарастает концентрация аллелей, которые сдвигают признак в направлении, нужном экспериментатору, но плохо «сочетаются» с аллелями других генов. Третья гипотеза принадлежит знаменитому английскому генетику К. Мазеру. Он предположил, что степень развития признака определяется совокупным эффектом большого числа локусов, аллели которых могут сдвигать величину признака в любом направлении. По мнению Мазера, слабые и сильные аллели (по эффекту на признак) разбросаны вдоль хромосомы случайно, а между ними находятся гены, влияющие на приспособленность и которые также представлены своими сильными и слабыми аллелями. При отборе, направленном на увеличение признака, сначала отбираются особи — носители хромосом с повышенным содержанием сильных аллелей. Затем в результате кроссоверной рекомбинации (т. е. обмена участками между гомологичными хромосомами) синтезируются хромосомы с еще большим числом сильных аллелей, однако новые (рекомбинантные) хромосомы могут нести увеличенное число аллелей, снижающих приспособленность. В итоге возникают генотипы, удовлетворяющие экспериментатора, но неудовлетворительные с точки зрения естественного отбора. Гипотеза неплохо объясняет длительные периоды «застоя» на кривых отбора как время ожидания синтеза рекомбинантных хромосом. Четвертая группа гипотез объясняет картину отбора мутациями. В связи с этим мы должны сначала кратко ознакомиться с основными открытиями Мукаи. В начале 60-х годов этот японский генетик приступил к серии экспериментов, продолжающихся и по сей день. На начальном этапе своих исследований Мукаи попытался оценить интенсивность и характер мутационного давления на гены, ответственные за жизнеспособность мух в лабораторных условиях. Для этого была заложена серия линий, причем геном каждой из них содержал копию одного вполне конкретного варианта второй хромосомы. В каждом поколении из хромосомного фонда линии случайно извлекался один экземпляр второй хромосомы и размножался. Заметим, что выбираемая хромосома передавалась от поколения к поколению без оценки ее влияния на приспособленность, т. е. без отбора. Кроме того, генетическая схема опыта позволяла на любом поколении сопоставить жизнеспособность мух, гомо- и гетерозиготных по данной хромосоме. У гомозигот оба экземпляра второй хромосомы были идентичными, тогда как у гетерозигот они происходили из разных линий. В начале опыта гомозиготы и гетерозиготы практически не различались по жизнеспособности, а затем во всех линиях начинался процесс постепенного «хирения» гомозигот, что можно объяснить накоплением второй хромосомой спонтанных мутаций с вредящим эффектом. Естественно, этот процесс шел во всех линиях совершенно независимо. Иногда случайно извлеченная хромосома счастливо избегала мутационных повреждений, в других случаях ей «везло» меньше. В некоторых случаях такая хромосома несла леталь, эти линии учитывались отдельно. Усреднив полученные данные по всем линиям (свободным от леталей), можно было наблюдать, как падает жизнеспособность у гомозигот по хромосоме, фактически выведенной из-под контроля очищающего отбора. К 60-му поколению она снизилась примерно в 2 раза. Мукаи установил, что это вызвано накоплением полудоминантных мутаций, каждая из которых снижает жизнеспособность мух всего на 2–4 %. Скорость их появления составляет 0,4 мутации на геном гаметы за генерацию. Такие мутации возникают в геноме в 20–30 раз чаще рецессивных с сильным повреждающим эффектом (леталей и полулеталей). Вернемся к искусственному отбору. В его ходе некоторый признак, обычно второстепенный для выживания (по крайней мере в лабораторных условиях) становится главным при разрешении вопроса жизни или смерти особи. Цель селекционера — сдвинуть в определенном направлении ценный, с его точки зрения, признак. Стремясь к этой цели, человек совершенно бессознательно способствует фиксации аллелей, благоприятствующих тому или иному уровню развития признака. Но локусы находятся в хромосомах, поэтому фиксация определенных аллелей — это фиксация довольно протяженных сегментов совершенно конкретных вариантов хромосом. Поддерживая в популяции высокий уровень благоприятных аллелей, селекционер фактически выводит из-под контроля естественного отбора сцепленные с ними хромосомные сегменты. Поэтому в них могут беспрепятственно накапливаться мутации, снижающие жизнеспособность и плодовитость. В природной обстановке очищающий отбор удалял бы эти мутации, но находящиеся рядом ценные для человека аллели их сохраняют. Поэтому, несмотря на прогрессивный однонаправленный сдвиг признака, в популяции наблюдается снижение главных показателей приспособленности. Возникает стандартная ситуация — цели человека входят в противоречие с естественным порядком вещей. В этих условиях определенную «помощь» популяции может принести кроссоверная рекомбинация, способная отделить аллели, ценные с точки зрения человека, от аллелей, вредных с точки зрения естественного отбора. Анализ генофонда популяций в ходе длительной лабораторной селекции показывает, что в нем действительно накапливаются мутации, снижающие главные компоненты приспособленности. В то же время, в процессе селекции появляются и благоприятные мутации, которые и позволяют в конечном счете преодолеть негативные эффекты отбора. Доместикация К настоящему времени накоплен довольно обширный материал (исторический и археологический), который позволяет проследить за изменением строения животных и растений при их доместикаций и в ходе дальнейшей культурной эволюции. Лучше всего этот материал освещает процесс доместикации в одном из ее главных очагов — регионе, история которого обычно излагается в разделе «Древний Восток». Этот географический район включает в себя обширную зону, в виде полумесяца охватившую восточную часть Средиземного моря, и две примыкающие к ней полосы. Одна — более широкая — протянулась от истоков Тигра и Евфрата вдоль западных склонов гор Загроса до Персидского залива, другая, узкая, вдоль Нила. На территории Древнего Востока примерно 9 тысяч лет назад произошло величайшее в истории человечества событие — переход от присваивающего хозяйства к производящему. Именно здесь человек одомашнил несколько видов животных и растений, которые и по сей день являются основными источниками его пищи. Не следует полагать, что человек сразу же после одомашнивания животных приступил к отбору на улучшение их хозяйственно ценных признаков. Все было гораздо прозаичнее — в течение 3–4 тысяч лет животных скорее всего съедали без всякой браковки. Такое положение вещей, вероятно, сохранялось примерно до момента возникновения первых цивилизаций в Междуречье и долине Нила. Обширные храмовые и царские хозяйства создали основу для возникновения отраслей экономики, ориентированных на производство мяса, молока, шерсти. С данного момента можно говорить о начале сознательной селекции, которая привела к созданию первых пород домашних животных. Переход к этой стадии сопровождался существенными изменениями морфологии, что запечатлено древними художниками Шумера и Египта. Можно проследить, как бараны теряют свою роскошную гриву и винтообразные рога, а их уши повисают. Любопытно, что примерно в это же время повисают уши и у коз, а у собак, быков и мелкого рогатого скота появляются пегости. Несколько позже громадные рога быков заменяются рожками шортгорнов. Все эти события датируются примерно вторым тысячелетием до нашей эры. Странный параллелизм в появлении признаков, отметаемых в природной обстановке очищающим отбором, можно приписать негативному эффекту искусственного отбора. Выше мы отмечали, что этот эффект можно в какой-то степени уменьшить синтезом новых, рекомбинантных хромосом, соединяющих аллели, полезные для человека, с аллелями, повышающими уровень главных компонент приспособленности. В таком случае искусственный отбор мог бы привести к повышению вероятности кроссоверного обмена и к более равномерному распределению этой вероятности вдоль хромосом. Недавно обнаружено, что уровень кроссоверной рекомбинации у домашних животных действительно гораздо выше, чем у диких. История домашних животных демонстрирует не только мощь искусственного отбора, но не в меньшей мере и процесс деградации структур, функция которых в новых условиях оказалась избыточной. Домашние животные сами не заняты поиском пастбищ и защищены человеком от хищников. Следовательно, по сравнению с дикими предками их существование в меньшей степени зависит от скорости и глубины обработки внешних сигналов. Все это означает снижение давления очищающего отбора, следящего за работой органов чувств и центральной нервной системы. В итоге у всех домашних животных по сравнению с их дикими сородичами зарегистрировано уменьшение объема головного мозга на 15–30 %. У многих видов уменьшен диаметр глазниц. Как правило, абсолютные размеры животных также снижены. Степень деградации соматических структур у крупного рогатого скота и лошадей достигла максимума в Центральной Европе к концу средних веков. Современные высокопродуктивные породы получены на основе фактически карликовых форм, оставшихся от средневековья. Исключение составляют скаковые лошади и некоторые породы собак, при выведении которых человек вел отбор на максимальную скорость бега. В этом случае цели человека не слишком расходились с обычными «целями» естественного отбора. В итоге деградация лошадей (прежде всего, уменьшение их размеров) связана обратной зависимостью с уровнем развития государства и его военных институтов. В частности, с падением Рима начался процесс резкого измельчания лошадей. Попробуем просуммировать некоторые особенности, отличающие домашних млекопитающих от их диких предков. Они включают в себя изменение характера окраски шкуры (прежде всего многочисленные пегости); потерю или редукцию рогов; уменьшение числа хвостовых позвонков; уменьшение размеров тела и уменьшение величины головного мозга и органов, связанных с восприятием внешних сигналов; вислоухость (т. е. неспособность двигать ушами). Все эти изменения свидетельствуют о деградации функций, повышающих выживание в естественной (дикой) обстановке. Пожалуй, только функция размножения была усилена в ходе доместикации, что и не удивительно, так как, стремясь увеличить продуктивность животных, человек автоматически отбирал особей, приносящих наибольшее число потомков. Итак, сходный (конвергентный) характер морфологических изменений при доместикации эволюционно неродственных видов скорее всего обусловлен снижением давления очищающего отбора, чему в немалой степени мог способствовать и интенсивный искусственный отбор, направленный на улучшение хозяйственно ценных признаков. Однако существует еще одно объяснение этой конвергенции. Советский генетик Д.К. Беляев предположил, что главная роль в процессе одомашнивания принадлежит отбору животных на ручной (неагрессивный) тип поведения. В ходе долговременного отбора на ручной тип поведения у серебристо-черных лисиц Д.К. Беляев с сотрудниками обнаружили немало наследуемых морфологических отклонений, вполне типичных для процесса одомашнивания. По мнению ученого, главная особенность отбора на поведение состоит в том, что он ведет к изменению гормонального статуса животных, а это, в свою очередь, может явиться причиной глубокой и направленной реорганизации онтогенеза. Постулируется, что изменение уровня гормонов может привести к наследуемому изменению активности ряда генов. Набор этих генов не вполне случайный, что и объясняет параллелизм в появлении аномальных форм у разных видов в процессе доместикации. Заметим, однако, что данная концепция оставляет без ответа многие вопросы. Получается, что сдвиг гормонального статуса взрослого организма может вызвать направленное изменение активности вполне определенных генов в клетках зародышевого пути, причем это измененное состояние генов способно сохраняться у потомков. Тем самым фактически предполагается превращение средовой (модификационной) изменчивости в наследственную. Отбор и интеграция процесса развития В ходе искусственного отбора происходит глубокая перестройка генофонда популяции, поэтому не следует думать, что из всех предложенных объяснений сопротивления популяции отбору верно только одно. Хотя мы и далеки от мысли, что все существующее разумно, но если природная популяция не гибнет в течение тысяч поколений, то, по-видимому, организация ее генофонда не так уж далека от оптимальной. Популяция не в силах полностью оградить себя от таких факторов, как мутационное давление, генный дрейф или генный поток, поэтому она всегда отягощена различными формами генетического груза. В то же время постоянное действие естественного отбора направлено на сохранение максимального в данных условиях уровня приспособленности. Для достижения этой цели популяция использует все доступные ей средства: от изменения способа размножения и уровня кроссоверной рекомбинации до изменения схемы индивидуального развития. Выше мы только вскользь упомянули о гипотезе С. Райта, которая объясняет сопротивление популяции действию отбора тонкой «подогнанностью» друг к другу аллелей разных локусов. Такая взаимозависимость эффектов генов скорее всего должна проявляться в процессе индивидуального развития организма. Рассмотрим абстрактную схему формирования взрослых структур А — F (рис. 15). За каждой из них стоит ряд зародышевых структур, нередко взаимодействующих друг с другом. На схеме эти зачатки условно разделены на два класса: поздние М — Р и ранние W— Y. Формально зиготу тоже можно считать зачатком (Z), общим для всех взрослых и эмбриональных структур организма. Каждая сплошная стрелка на схеме олицетворяет собой все многообразие генов, ответственных за развитие более раннего зачатка в более поздний. Эти гены можно символизировать сочетанием двух прописных букв — первая относится к более раннему зачатку, вторая — к более позднему. Взаимодействия типа эмбриональной индукции обозначены на схеме горизонтальными пунктирными линиями. Представим себе, что селекционер стремится изменить вполне конкретную взрослую структуру. Ясно, что для этого он должен в каких-то генах, скрытых за стрелками, ведущими к данной структуре, зафиксировать новые (благоприятные с точки зрения селекционера) аллели. Из рис. 15 видно, что для изменения структуры А можно замещать аллели в генах ZW, WM и МА.
 Рис. 15. Схема индивидуального развития структур организма.
Z — зигота; W—Y — ранние зачатки; M — R — поздние зачатки; А — F — взрослые структуры. Штриховой линией отмечена эмбриональная индукция.
Рис. 15. Схема индивидуального развития структур организма.
Z — зигота; W—Y — ранние зачатки; M — R — поздние зачатки; А — F — взрослые структуры. Штриховой линией отмечена эмбриональная индукция.
Обратим внимание на путь формирования структуры В. Отметим, что из позднего зачатка N кроме нее возникает еще одна взрослая структура С. Если эта последняя прямо связана с основными показателями приспособленности, то ее развитие должно быть оптимизировано естественным отбором. Для изменения структуры В можно замещать аллели в любых локусах, стоящих за стрелками, ведущими к ней. Однако такие замещения в ранних генах кроме изменения структуры В могут затронуть еще одну структуру — С, а это уже может снизить уровень главных показателей приспособленности. Чтобы скомпенсировать данный негативный эффект, следовало бы структуре С возвратить ее прежнее значение, зафиксировав для этого новый аллель по одному из поздних локусов NC, т. е. в данном случае для адаптивного сдвига нужно зафиксировать два новых аллеля. Обобщая этот пример, можно показать, что чем больше нитей связывает развитие какой-то структуры с другими, тем больше новых аллелей нужно зафиксировать для достижения одного и того же сдвига структуры у взрослой особи. Выходит, что отбор в основном должен оперировать с генами, которые управляют завершающими стадиями развития организма, тогда как гены, контролирующие ранние стадии, фактически не допускают изменений. Это хорошо объясняет сохранение основного плана строения животного в ходе эволюции. В то же время, чем раньше обособляется зачаток, тем выше шансы на эволюционный сдвиг у развивающейся из этого зачатка структуры. Любопытно, что у растений негативные эффекты искусственного отбора выражены гораздо меньше, чем у животных. Особенно впечатляюще выглядят результаты знаменитого Иллинойсского эксперимента. Группа американских селекционеров в самом конце XIX века приступила к отбору, направленному на изменение ряда количественных признаков у кукурузы. Этот опыт продолжается до сих пор. На рис. 16 показано, как изменялось в ходе 76 поколений отбора содержание масла в семенах. Как видно, ответ на отбор может сохраняться в течение весьма продолжительного периода.
 Рис. 16. Отбор на изменение содержания масла в семенах кукурузы (по: [Dudley, 1977]).
Рис. 16. Отбор на изменение содержания масла в семенах кукурузы (по: [Dudley, 1977]).
Эту особенность растений можно объяснить с двух позиций. Во-первых, в отличие от животных в жизненном цикле растений есть стадия гаплоидного организма — гаметофита. Всем известная цветочная пыльца является скоплением мужских гаметофитов. Биомасса пыльцевого зерна — продукт деятельности гаплоидного генома одного ядра. Этот же геном обеспечивает превращение пыльцевого зерна в трубку, прорастающую сквозь пестик до завязи. По этой трубке в зародышевый мешок доставляются мужские гаметы. Избыток пыльцевых зерен создает условия для соревнования гаметофитов по оплодотворяющей способности, т. е. на стадии мужского гаметофита идет как бы проверка большого числа генов на качество их работы. Это можно рассматривать как приспособление для снижения мутационного генетического груза. Не исключено, что еще одной причиной слабого сопротивления растений давлению отбора является довольно низкая степень интеграции их индивидуального развития. Об этом может свидетельствовать, например, отсутствие каких-либо нарушений в развитии гибридов, полученных при соединении весьма отдаленных геномов. Хорошо известна мощность и высокая фертильность амфидиплоидов, у которых совмещены геномы растений, принадлежащих разным родам. Достаточно вспомнить о мягкой пшенице, совмещающей геномы твердой пшеницы и эгилопса, или об искусственно полученном пшенично-ржаном гибриде — тритикале. Ограничения на скорость эволюции Поставим в центр нашего рассмотрения вид — главное действующее лицо в макроэволюционной драме. Его жизнь протекает в очень сложной экологической обстановке. Образно говоря, ему постоянно приходится заботиться о двух вещах: о возобновлении пищевых ресурсов и о защите своих особей от истребления. Пищевые и прочие ресурсы вид, как правило, делит с другими видами-конкурентами, а жизни его особей угрожают многочисленные хищники. К тому же пищей для рассматриваемого вида обычно служат живые организмы, все время пытающиеся поднять свою приспособленность, вырабатывая какие-то специальные средства защиты и сокращая тем самым ресурсы вида. Наконец, над всей этой борьбой стоят геологические и географические факторы. Изолирующие барьеры возникают и исчезают, климат меняется, происходят засоление и опреснение водоемов и т. д. Все это ухудшает условия жизни вида. Собственно говоря, возникает естественный вопрос, как вообще можно существовать в таких экстремальных условиях, да еще на протяжении миллионов лет. Этот вопрос несколько сродни другому: как каждый отдельный человек умудряется дожить до седин, если, оглядываясь на прожитую жизнь, он может насчитать до десятка случаев, когда был на волосок от смерти. Тем не менее довольно высокая средняя продолжительность жизни явно свидетельствует о том, что риск погибнуть во цвете лет не так уж велик. Жизнь вида представляется нам такой ненадежной потому, что мы несколько сгустили краски, просуммировав все факторы опасности. Реальная ситуация обычно не так уж страшна, так как давление неблагоприятных факторов и главное скорость нарастания этого давления не слишком велики. Наконец, вид, разобщенный сложной географической средой на огромное число изолированных популяций, не может быть всюду застигнут врасплох действием неблагоприятного фактора. Какая-то его часть может спастись и после разрушения изолирующих барьеров занять свой прежний ареал. Правда, такой способ выживания эволюцией (тем более прогрессивной) не назовешь, но в эффективности ему не откажешь. Теперь представим себе ситуацию, когда виду, образно выражаясь, отступать некуда, поскольку он представлен только одной компактной популяцией, и на него надвигается неблагоприятный фактор: вид-конкурент или вид-хищник, или просто ухудшается климат. В данной ситуации выживание вида определяется исключительно скоростью преобразования его наследственной информации. Старых «знаний» о среде ему теперь недостаточно, их необходимо подновить. На языке популяционной генетики такое подновление означает замещение аллелей в нескольких генных локусах. Хорошо, если эти аллели в генетическом фонде вида уже имеются, притом в приличных концентрациях. В популяциях, насчитывающих миллион особей и больше, величина 2NV обычно превосходит единицу, т. е. благоприятные аллели имеются всегда, хотя их частоты должны быть очень низки (см. уравнение (3.31)). Естественный отбор в этих условиях мало эффективен, поэтому, несмотря на повторное появление, большинство благоприятных мутаций популяция должна терять. При наличии изолирующих барьеров большое значение приобретает генный дрейф, который может в некоторых субпопуляциях быстро поднять частоту благоприятного аллеля и сделать это совершенно «бесплатно», т. е. без селективной гибели менее приспособленных. Правда, вероятность такого везения невелика, но, скорее всего, оно является необходимым условием для начала действия движущего отбора — по существу, единственного фактора, способного быстро завершить фиксацию благоприятного аллеля. Однако прямое воздействие отбора не может сдвинуть значение количественного признака на любой наперед заданный процент. В этом отношении материал организмов напоминает по своим свойствам совсем не пластилин, а твердое на ощупь аморфное тело, наподобие стекла. Такие тела, с точки зрения физика, являются жидкостями с очень большим коэффициентом вязкости, поэтому под влияниям внешнего воздействия они могут течь, т. е. изменять свою форму в любом направлении. В отличие от них настоящие твердые тела сопротивляются попыткам изменить их форму. У обычных жидкостей, вроде воды, такое сопротивление практически отсутствует, поскольку они способны чрезвычайно быстро (за 10-12—10-10 секунд) перестроить под влиянием внешнего воздействия свою молекулярную структуру. У очень вязких жидкостей время, необходимое для такой перестройки, на много порядков больше. По отношению к быстро меняющимся воздействиям очень вязкие жидкости ведут себя как хрупкие тела, ломаясь даже при небольших деформациях. Чтобы заставить аморфные тела течь, нужно прикладывать к ним не очень большие, но весьма длительные нагрузки. Время воздействия на такую «твердую» жидкость должно быть тем больше, чем выше коэффициент ее вязкости. В случае изменения морфологии под действием отбора роль этого коэффициента выполняют прежде всего межгенные взаимодействия, контролирующие индивидуальное развитие организма. Если давление отбора слишком велико, то система межгенных взаимодействий не успевает перестроиться, и популяция вымирает. При полной (абсолютной) интеграции онтогенеза все мутации стали бы неблагоприятными, так как любая из них сказалась бы отрицательно на большинстве основных функций организма. В связи с этим легкость перестройки генетической программы развития определяется не столько мутабильностью генов, сколько уровнем разобщенности (автономности) процессов развития зачатков рабочих структур, обслуживающих основные функции. Прекрасным примером такой автономности служит очень раннее обособление у животных зародышевого пути, ведущего к созданию главных структур функции воспроизведения, — гамет. Мы видим, что степень разобщенности путей развития рабочих структур может сказаться на скорости фиксации благоприятных мутаций, т. е. на степени эволюционной пластичности видов. Следовательно, эта пластичность в определенной мере может зависеть от информационного содержания генома и, в частности, от генетического контроля процессов индивидуального развития. Другое ограничение на скорость адаптивной эволюции связано с мутационным грузом. Интенсивный движущий отбор, быстро фиксируя благоприятные мутации, фиксирует и сегменты хромосом, в которых эти мутации расположены. Тем самым значительная часть генома выходит из-под контроля очищающего отбора, что ведет к быстрой аккумуляции в ней слегка вредящих мутаций. Очевидно, что ответ на отбор прекратится, когда позитивный эффект благоприятных мутаций уравновесится негативным эффектом вредящих. В связи с этим скорость эволюции попадает в зависимость от скорости освобождения хромосом от мутаций с отрицательным эффектом на главные показатели приспособленности. Наконец, очевидно, что скорость фиксации благоприятных мутаций лимитируется скоростью их появления. Последняя же в соответствии с (3.32) определяется «выходом» таких мутаций, т. е. их долей среди всех мутаций, возникающих в эволюционирующей популяции за одно поколение.
Макроэволюция
Глава 5. Мощность и сложность
Если бы возможно было показать, что существует сложный орган, который не мог образоваться путем многочисленных, последовательных, незначительных изменений, моя теория потерпела бы полное крушение. Но я не могу найти такого случая.Филетическая эволюция В ходе экологической дифференциации отщепившаяся от вида популяция может со временем стать новым видом. Заметим, что при этом может продолжать жить и исходный (материнский) вид. Иная судьба складывается у вида, когда среда его обитания медленно, но неуклонно ухудшается. Обычно такое ухудшение касается какого-то одного жизненно важного параметра. Само собой разумеется, что любой вид существует лишь в пределах вполне конкретного диапазона изменения такого параметра и вымирает при выходе среды за эти пределы. Следовательно, выживание при постепенном ухудшении среды должно быть связано с особой формой видообразования, при которой материнский вид прекращает свое существование. Такое видообразование получило название филетического. Однако возник ли новый вид, отщепившись от материнского, или же он является его филетическим потомком, во всех случаях у любого вида был вид-предок, а этот последний также имел предка и т. д. Таким образом, каждый вид многоклеточных организмов представляет собой завершающее звено длинной цепи видов-предков (филетической линии вида), начало которой теряется в глубинах докембрия. Заметим, что филетическая цепь каждого вида начиналась в совершенно других, по существу, инопланетных условиях. Конечно, это верно и для нас — людей. Вообразим, что мы опускаемся по нашей филетической цепи в глубь времен. Сначала пройдем через звенья сухопутных форм млекопитающих и рептилий, войдем в контакт с водой на стадии амфибий, погрузимся под воду на участках, принадлежащих костным рыбам и их круглоротым предкам, опустимся на морское дно, проходя стадии придонных фильтраторов, подобных ланцетнику… А глубже, где-то во мраке времен, мы минуем звенья, занятые червями и медузоидами и, наконец, достигнем самого протяженного участка, принадлежащего царству одноклеточных. На Земле в настоящее время существуют миллионы видов животных и растений, значит, существует столько же непрерывающихся филетических линий. Это ли не свидетельство силы жизни! К сожалению, лишь малая часть линий смогла дожить до наших дней. Если бы нам представилась фантастическая возможность подняться из прошлого к настоящему, то в большинстве случаев достичь современности нам бы не удалось. Сплошь да рядом мы натыкались бы на конечные звенья, т. е. на виды, которые вымерли. Кроме того, поднимаясь по филетическим цепям, мы бы очень часто встречались с их ветвлением, где перед нами вставала бы проблема выбора маршрута. Такие развилки проще всего трактовать как завершение экологической дифференциации изолированных популяций. Вновь возникшие ветви могут опять ветвиться, преобразуя филетическую линию в филетический пучок. Судьба филетических линий такого пучка нередко складывается однотипно — на каком-то весьма кратком, с геологической точки зрения, отрезке времени все они могут вымереть. И наоборот, отдельная линия пучка может испытать бурное ветвление, что приведет к возникновению нового пучка — дочернего по отношению к исходному. Процесс расщепления филетических пучков может продолжаться и далее. В итоге, возникает иерархия пучков разного калибра, образовавшихся из одной филетической линии. Такое монофилетическое собрание пучков будем называть филетической группой. Филетические линии и их объединения отличаются от иерархически построенных групп систематиков тем, что здесь в явной форме присутствует время. Проведя как бы временной срез через любую филетическую группу, мы получим таксоны систематиков. Филетические линии превратятся в виды, а пучки и их группы — в таксоны более высокого ранга: роды, семейства, отряды и т. д. Оперируя с филетическими линиями и их пучками, мы фиксируем свое внимание на том общем (гомологичном), что сохраняют особи разных видов на протяжении миллионов лет. При таком макроэволюционном подходе отдельные виды (вместе с царящими в них микроэволюционными процессами) представляются нам неделимыми атомами, а наш основной интерес прикован к тому, что происходит с гомологичными структурами (как морфологическими, так и молекулярными) в течение долгой жизни филетических линий. Говоря о макроэволюции, мы пользуемся широко принятой аналогией между филетической группой и деревом. Развивая ее дальше, можно сравнить филетическое видообразование с ростом ветвей. Правда, в отличие от обычного дерева за этим филогенетическим древом следит «садовник», который время от времени подстригает побеги, лишая ветви способности к дальнейшему росту. Этот садовник в своей работе придерживается некоторых правил: во-первых, он подстригает только ветви, расположенные на максимальной высоте, и, во-вторых, этой операции нередко подвергаются все побеги одной крупной ветви, включающей в себя множество более мелких ветвей и веточек. Ясно, что в качестве садовника выступают изменения среды, адаптироваться к которым некоторые филетические линии не смогли, в то время как прирост ветвей и образование боковых побегов — это всегда успешно завершенный процесс адаптации к новым (обычно неблагоприятным) условиям среды. Странное пристрастие «садовника» к некоторым крупным ветвям наводит на мысль, что все виды монофилетического таксона наследуют от своего общего предка нечто, определяющее их способность к филетическому выживанию в изменяющейся среде. Массовые вымирания давно волнуют биологов. Шутка ли, когда в считанные миллионы лет гибнут пучки, представленные тысячами филетических линий. Достаточно вспомнить о гибели наиболее продвинутых по пути прогресса рептилий в конце мелового периода, причем таких разных, как динозавры на суше, птерозавры в воздухе и ихтиозавры в воде. В конце концов гибель всегда можно как-то объяснить (взрыв сверхновой, падение астероида, массовые извержения вулканов и пр.); непонятно только, почему одни гибнут, а другие, вроде бы не лучше и не хуже, продолжают процветать. Если расцвет млекопитающих и птиц можно объяснить их теплокровностью, то почему вместе с динозаврами не погибли ящерицы и змеи? Почему печальная участь аммоноидей не постигла головоногих моллюсков с внутренней раковиной? Палеонтологи разделяют фанерозой (см. таблицу) на 80—100 геологических веков (по 6–7 миллионов лет на век), каждый из которых существенно отличается от соседних по видовому и даже родовому составу организмов. Смена фауны на границе веков охватывала огромные территории и нередко сопровождалась массовой гибелью многих филетических пучков. Вместе с тем, несмотря на сравнительно небольшую продолжительность геологического века, этого времени нередко оказывалось достаточно для того, чтобы расщепившиеся популяции достигли межродового ранга. Иногда темп морфологической эволюции бывал еще выше. Например, в начале фаменского века (завершившего девон) возникли климении — очень своеобразная группа аммоноидей с несколько измененным планом строения (у них сифон — важная деталь гидростатического аппарата — вдруг переместился с дорсальной части раковинной трубки на вентральную). Таксономическая эволюция климений протекала чрезвычайно бурно. За время одного геологического века они успели породить несколько семейств, насчитывавших десятки родов. Крупные преобразования затронули многие признаки. Некоторые из морфологических достижений климений, например треугольное завивание раковины, так и не были воспроизведены другими аммоноидеями, хотя им судьба даровала для экспериментирования более 300 миллионов лет. Тем не менее, несмотря на свое поразительное разнообразие, все климении вымерли в конце фаменского века. Правда, ради справедливости следует отметить, что изменение среды, отделившее девон от карбона, носило характер глобальной крупномасштабной катастрофы. Другие отряды аммоноидей преодолели это испытание с большим трудом. Адаптируемость Итак, чем же определяется эволюционная судьба филетических групп? Почему одни из них без труда преодолевают все катастрофы, выпавшие на их судьбу за сотни миллионов лет,тоща как другие, эволюционируя в одной и той же адаптивной зоне, после вспышек ветвления внезапно исчезают. Конечно, все это многообразие эволюционных судеб можно было бы объяснить игрой случая, однако именно с точки зрения статистики трудно поверить в синхронное вымирание сотен филетических линий, собранных в крупные филетические пучки. Если вероятность вымирания каждой линии одна и та же, то медленные изменения среды, описанные в последнем параграфе гл.3, не должны полностью истреблять большие филетические группы. Предположим, что вероятность вымирания определяется одним внутренним свойством линии — генетически определяемой способностью преодолевать долговременные, неблагоприятные для нее изменения среды. Назовем это свойство адаптируемостью. Как закодированное в геноме оно должно наследоваться при видообразовании, т. е. передаваться (с легкими изменениями) от вида-предка к виду-потомку. Следовательно, виды-соседи по филетической цепи и даже виды из разных линий одного филетического пучка должны обладать близкими значениями адаптируемости. Виды с высоким значением адаптируемости имеют больше шансов преодолеть неблагоприятные изменения среды, поэтому средняя адаптируемость у видов-потомков должна быть несколько выше, чем у видов-предков. За время существования многоклеточных (700–800 миллионов лет) произошло около ста довольно крупных изменений фауны, которые могут служить границами ярусов — стратиграфических аналогов геологических веков. Можно предположить, что филетические линии, выдержавшие в конце концов все эти испытания, отличаются от вымерших повышенной скоростью преобразования наследственной информации. На филетический пучок можно взглянуть как на популяцию бесполых организмов. В такой популяции каждая особь (отдельный вид) через фиксированный промежуток времени (равный продолжительности жизни вида) или гибнет, или оставляет потомков (дочерние виды). При этом адаптируемость вида — это приспособленность отдельной особи в такой популяции, т. е. адаптируемость можно определить как вклад данного вида в филетический пучок на следующем отрезке макроэволюционного времени. Заметим, что длительность такого отрезка соизмерима с миллионом лет. Как показал в 1955 г. американский генетик Дж. Кроу, в бесполой популяции идет постоянное возрастание средней приспособленности. При этом скорость данного процесса в точности равна популяционной дисперсии по приспособленности. Поскольку дисперсия (как квадрат сигмы) не может быть отрицательной величиной, то средняя адаптируемость филетического пучка в ходе эволюции может только возрастать. Не является ли это свидетельством эволюционного прогресса? Эволюционная адаптация к новой среде предполагает фиксацию серии благоприятных мутаций. В соответствии с уравнением (3.31) скорость этого процесса прямо зависит от размера популяции, скорости появления благоприятных мутаций и степени их благоприятности. Мы определили адаптируемость как внутреннее, т. е. заключенное в геномах особей, свойство вида. Из перечисленных выше факторов лучше всего ей соответствует скорость появления благоприятных мутаций. Но процесс мутирования слеп, поэтому из всех возникающих мутаций лишь небольшая их часть может быть благоприятной в данной конкретной (неблагоприятной для популяции) обстановке. Следовательно, для повышения адаптируемости было бы неплохо увеличить скорость появления мутаций, способных компенсировать изменения среды, наиболее частые в экологическом пространстве вида. Поскольку мы ввели адаптируемость как неотъемлемое свойство генома типичной особи вида, то тем самым признали, что могут быть мутации как уменьшающие, так и увеличивающие адаптируемость. Фактически мы предположили, что могут существовать мутации, изменяющие скорость появления благоприятных мутаций. На первый взгляд кажется, что для этого достаточно просто повысить мутабильность среднего гена, однако такое решение вопроса весьма сомнительно. Ведь рост мутабильности автоматически означает увеличение генетического груза, т. е. падение уровня приспособленности популяции. Наиболее перспективно выглядят мутации, повышающие долю благоприятных среди всех мутаций, возникающих в геноме за поколение. Если такие мутации, какими бы фантастичными они ни казались, действительно существуют, то они должны накапливаться в генофондах видов при браковке филетических линий по адаптируемости. В настоящее время существуют две точки зрения на филетическое видообразование. Одна традиционная, градуалистская, впервые высказанная Дарвином, предполагает, что нет четкой границы, где один вид сменяет другой, поэтому сдвиг фенотипа средней особи идет постоянно, хотя в его скорости возможны значительные колебания. Согласно другой, высказанной С. Гоулдом и Н. Элдриджем в 1972 г., вид в течение миллионов лет не изменяется, находясь в состоянии морфологического «стазиса». Затем за относительно короткий промежуток времени происходит его превращение в новый вид — видовая трансформация. Хотя этот «краткий» миг может длиться и несколько десятков тысяч лет, все равно его длительность ничтожна в сравнении с миллионами лет стазиса. Если верна градуалистская модель, то процесс адаптации к новым условиям длится миллионы лет, и все это время сохраняются условия для фиксации мутаций, увеличивающих адаптируемость. Пунктуалистская модель предполагает, что отбор в пользу таких мутаций действует наверняка только в период видовой трансформации, и ценные для этого момента аллели могут быть утрачены во время стазиса, если они не вносят вклад в относительную приспособленность особей. Следовательно, аллели, благоприятные для филетического выживания вида, должны быть благоприятными и в микроэволюционном смысле, т. е. повышать уровень относительной приспособленности их носителей. Заметим, что, введя представление об адаптируемости, мы получили, как следствие, макроэволюционную тенденцию к ее росту. Эту тенденцию можно было бы интерпретировать как накопление эволюционирующей филетической группой информации о неблагоприятных факторах, действующих в ее экологическом пространстве (адаптивной зоне). Как подойти к анализу подобного рода информации, вероятно, запечатленной геномом? Скорее всего такая генетическая запись должна каким-то образом отразиться на фенотипе особей. Поэтому и тенденция к росту адаптируемости в ходе эволюции филетической группы могла бы выглядеть как долговременная тенденция к изменению в определенном направлении морфологии особей ее видов. В связи с этим встает вопрос, не являются ли хорошо известные палеонтологам долговременные тенденции отражением процесса возрастания адаптируемости. Макроэволюционные тенденции Крупнейший вклад палеонтологии в наши знания об эволюции живых организмов состоит в обнаружении во многих филетических группах особых очень долговременных тенденций к сдвигу в одном направлении некоторых количественных признаков. Пожалуй, самой известной из них является тенденция к увеличению размеров индивидов — так называемое правило Копа. Значительный эмпирический материал свидетельствует, что обычно зоологические таксоны крупных животных происходят от видов с относительно небольшими особями. Между прочим, род Homo происходит от гораздо более мелких обезьян. Э. Коп объяснил свое правило как классический ламаркист (кстати, основоположник психоламаркизма). Он считал, что в основе открытой им тенденции лежит пресловутая «ростовая сила», стремящаяся под действием психики животного увеличить его размеры. Такой филогенетический рост тела протекает беспрепятственно в периоды геологического затишья и сопровождается разрастанием многих морфологических структур. Тенденции противостоит процесс вымирания гигантов и монстров при резких сменах климата.Ч. Дарвин
 Рис. 17. Изменение видовых распределений брахиопод (а) и аммонитин (б) по размеру раковины в историческом развитии (по: [Stanley, 1973]).
Рис. 17. Изменение видовых распределений брахиопод (а) и аммонитин (б) по размеру раковины в историческом развитии (по: [Stanley, 1973]).
Отметим одну особеннность коповского ламаркизма: внутренние стремления индивида снижают адаптируемость филетической линии и влекут последнюю к гибели. Любопытно, что включение в эволюционный сценарий особых роковых факторов — довольно обычное дело для теоретиков-эволюционистов. Достаточно вспомнить о трех таких драматических историях: 1) самоубийственной тенденции юрских устриц (грифей) к созданию нераскрывающихся раковинных створок; 2) инадаптивном сверхразрастании непомерно тяжелых рогов у ирландского оленя; 3) развитии сверхмощных клыков, затрудняющих кусание, у саблезубых кошек. Все эти линии на самом деле вымерли, что, конечно, не противоречит предположению об инадаптивном характере данных тенденций. Правда, вымирание филетических линий — дело обычное. Ведь вымерли же, например, все виды семейства Hominidae, за исключением Н. sapiens, и вообще филетический пучок высших приматов не выглядит преуспевающим. Не станем же мы говорить об инадаптивном сверхразрастании головного мозга у гоминид. Кроме того, отметим, что все отмеченные выше случаи «сверхразвития» рабочих структур противоречат принципу симморфоза, согласно которому ни один орган не бывает развит свыше потребности в нем в моменты максимальных нагрузок. Однако вернемся к правилу Копа. Проследим в какой-нибудь филетической группе за динамикой видовых распределений по величине особей. На рис. 17, а приведены распределения по диаметру раковины у брахиопод отряда Rynchonellida на трех этапах их исторического развития. Аналогичную картину демонстрируют и аммонитины юры (рис. 17, б). Первое, что бросается в глаза, — это увеличение размаха распределений, обусловленное прежде всего появлением видов с большими размерами особей. Кроме того, эти распределения асимметричны — с длинным «хвостом» в правой части. Очевидно, что среднее значение (центр тяжести) распределений также сдвигается с ходом времени вправо. Отметим, что виды с мелкими особями продолжали существовать всегда, хотя их доля постепенно убывала. Сходную картину можно наблюдать и для видовых распределений по признакам, величина которых хорошо коррелирует с массой тела. Чтобы понять это макроэволюционное явление, сначала рассмотрим, как вообще формируются распределения видов по количественным признакам. Отметим, что после перехода к логарифмической шкале измерения признака многие из этих распределений симметризуются, становясь близкими к нормальным. Межвидовые дистанции Мы на глаз, без труда различаем млекопитающих, принадлежащих разным родам. Например, волка и лисицу, рысь и леопарда, козу и овцу и т. д. Еще легче разобраться с представителями разных семейств, отрядов и других таксонов высокого ранга. Сортировка видов одного рода требует большего внимания, хотя в случае млекопитающих она обычно удается, вероятно, помогает собственная принадлежность к данному классу. Видимо, поэтому мы можем отличить друг от друга льва, тигра и леопарда, несмотря на то, что все они входят в один род Panthera. Кстати, отличить череп льва от черепа тигра под силу только специалисту, а гибрид этих самых сильных кошек — тигролев — вполне жизнеспособен. Куда труднее различить родственные виды у насекомых, особенно когда речь идет о так называемых видах-двойниках. Человек при оценке степени морфологического сходства пользуется присущей ему интуитивной способностью распознавать образы. «Ненаучность» такого метода очевидна. Первая вполне научная система организмов была создана К. Линнеем в 1735 г. Она основана на сопоставлении рядов четко регистрируемых морфологических признаков, которыми можно охарактеризовать типичную особь. Чем больше признаков совпадает в таких рядах, тем выше степень сходства сравниваемых видов. Чем меньше число совпадений, тем выше ранг таксономической дистанции между видами. Слабость этого подхода заключается в субъективном характере выбора признаков, положенных в основу системы. К сожалению, более объективные молекулярные методы обычно дают информацию только о времени эволюционной дивергенции видов. Дело в том, что удобные для регистрации различия в строении белков или нуклеиновых кислот чаще всего обусловлены нейтральными аллельными заменами, которые фиксируются с более или менее постоянной скоростью — скоростью мутирования (см. (3.26)). В настоящее время существует ряд способов измерения так называемой морфологической (фенотипической) дистанции между двумя группами особей. Несмотря на различия в математическом оформлении, все эти оценки являются сложным отражением того, во сколько раз (или на сколько процентов) отличаются значения одних и тех же признаков у сравниваемых групп особей. По-видимому, из-за этого оцененные разными способами значения морфологических дистанций неплохо коррелируют друг с другом. Многочисленные измерения фенотипических дистанций позволяют прийти к некоторым выводам, довольно существенным для нашей темы. Во-первых, для многих совсем не родственных видов животных средние расстояния между популяциями одного вида обычно весьма близки, и, во-вторых, для морфологических дистанций, отделяющих виды одного рода, также характерен свой средний уровень, который, как правило, в 1,5–2 раза превосходит межпопуляционный. Эти результаты наводят на мысль, что превращение популяций в виды сопряжено с достижением их особями вполне определенного значения относительного сдвига величины ряда количественных признаков. Что же происходит с видом в момент видообразования? Очевидно, приспособление к новым экологическим условиям. Хорошо известно, что для каждого вида ряд жизненно важных параметров среды не должен выходить за пределы некоторых критических значений. Так, для жизни рифообразующих коралловых полипов необходимо, чтобы температура воды не опускалась ниже 20,5 °C. Полный комплекс требований вида к параметрам среды может быть объединен понятием экологической ниши вида. Таким образом, каждому виду соответствует его экологическая ниша. В одной нише устойчиво может существовать только один вид (принцип Гаузе), если вдруг по какой-то причине в ней окажутся два, то со временем один из них уступит место другому, скорее всего, он вымрет. Филетическое выживание вида (видообразование) сопряжено с его переходом в соседнюю экологическую нишу, при этом же, очевидно, происходят сдвиги значений многих количественных признаков особей. Мы уже отмечали (см. параграф «Аллометрия» в гл.2), что сдвиги подобного рода обычно связаны с изменением абсолютных размеров структур и прежде всего с изменением средней массы животного. Знаменитый американский эколог Дж. Хатчинсон предположил, что для завершения экологической дифференциации средняя величина морфологического признака, непосредственно ответственного за адаптацию, должна измениться в какое-то фиксированное число раз. Оказалось, что довольно часто выполняется одно удивительное правило: массы особей родственных видов животных, обитающих в одной местности (т. е. в сходных экологических условиях), отличаются друг от друга в число раз, примерно кратное двум. Как будто для экологического разобщения особи таких видов должны различаться по массе примерно в 2 раза. Из этого правила Хатчинсона следует, что линейные размеры тела отличаются минимум в 21/3 ≈ 1,26 раза, или, грубо, на 30 %. Особенно четко это правило выполняется для размеров пищедобывающих структур животных. Прямые измерения показывают, что чаще всего линейные размеры этих структур (L) отличаются, как минимум, на 28 %. Отсюда вытекает, что размер L можно представить как некую минимальную величину Lmin, умноженную на произведение ряда сомножителей хi, близких к 1,28, т. е.
 (5.1)
После логарифмирования этого выражения имеем
(5.1)
После логарифмирования этого выражения имеем
 (5.2)
Заметим, что ∑lnxi представляет собой сумму близких по порядку независимо варьирующих величин, поэтому распределения большого числа видов по логарифму размеров или массы особей должны быть близкими к нормальным.
Формирование видовых распределений
Построим простейшую модель видовых распределений. Будем считать, что для экологической дифференциации видов их особи должны отличаться по размерам на 28 %, соответственно натуральный логарифм их размеров — приблизительно на 0,25. Разобьем горизонтальную ось, вдоль которой изменяется логарифм линейных размеров, на интервалы шириной 0,25. Пусть вид-основатель филетической группы по размеру своих особей находился в интервале х0. Через единичный отрезок времени (через один шаг модели) он произведет два дочерних вида, которые попадут в соседние интервалы х-1, и х1. Отметим, что интервал х0 остается занятым материнским видом. В следующий временной отрезок каждый из трех видов приступит к «завоеванию» своих соседних интервалов. Продолжая такой процесс, мы вскоре получим довольно гладкое симметричное (биномиальное) распределение, очень близкое к нормальному. С каждым шагом оно расширяется, его дисперсия увеличивается пропорционально возрасту таксона, а сигма следует за квадратным корнем из этого возраста. Однако центр тяжести распределения (его среднее значение) должен оставаться на месте, точно соответствуя величине признака у вида-основателя филетической группы — х0 (рис. 18, а).
Ясно, что такая модель не объясняет правила Копа. Начнем ее усложнять. Быть может, мы получим искомый сдвиг распределения вправо, если зафиксируем его левую границу, т. е. примем, что вид с величиной тела меньше некоторого критического значения х* просто не может существовать. Подобное соображение выглядит вполне реалистичным. Например, для очень мелких теплокровных животных, таких как колибри у птиц и землеройки у млекопитающих, дальнейшее уменьшение размеров создало бы серьезные проблемы с поддержанием температуры тела. По-видимому, колибри, питающиеся очень калорийным нектаром, уже достигли левой границы распределения птиц. Кстати, в покое они не в состоянии поддерживать постоянную температуру.
Пусть критическое значение признака соответствует интервалу x-8. Отсюда следует, что вид, занимающий этот самый левый интервал, может дать дочерний вид только с увеличенным размером особей, заселяя интервал х-7. С ходом времени мы получим распределение, которое будет весьма походить на предыдущее, отличаясь от него практически только отсутствием части, расположенной слева от «поглощающего экрана» (рис. 18, б). Естественно, с ходом времени среднее значение такого урезанного распределения будет смещаться вправо, а незначительным сдвигом его вершины (моды) можно пренебречь. Однако тщательный анализ реальных видовых распределений (подобных представленным на рис. 17) обнаруживает четкое смещение их вершины вправо, т. е. в сторону увеличения признака.
(5.2)
Заметим, что ∑lnxi представляет собой сумму близких по порядку независимо варьирующих величин, поэтому распределения большого числа видов по логарифму размеров или массы особей должны быть близкими к нормальным.
Формирование видовых распределений
Построим простейшую модель видовых распределений. Будем считать, что для экологической дифференциации видов их особи должны отличаться по размерам на 28 %, соответственно натуральный логарифм их размеров — приблизительно на 0,25. Разобьем горизонтальную ось, вдоль которой изменяется логарифм линейных размеров, на интервалы шириной 0,25. Пусть вид-основатель филетической группы по размеру своих особей находился в интервале х0. Через единичный отрезок времени (через один шаг модели) он произведет два дочерних вида, которые попадут в соседние интервалы х-1, и х1. Отметим, что интервал х0 остается занятым материнским видом. В следующий временной отрезок каждый из трех видов приступит к «завоеванию» своих соседних интервалов. Продолжая такой процесс, мы вскоре получим довольно гладкое симметричное (биномиальное) распределение, очень близкое к нормальному. С каждым шагом оно расширяется, его дисперсия увеличивается пропорционально возрасту таксона, а сигма следует за квадратным корнем из этого возраста. Однако центр тяжести распределения (его среднее значение) должен оставаться на месте, точно соответствуя величине признака у вида-основателя филетической группы — х0 (рис. 18, а).
Ясно, что такая модель не объясняет правила Копа. Начнем ее усложнять. Быть может, мы получим искомый сдвиг распределения вправо, если зафиксируем его левую границу, т. е. примем, что вид с величиной тела меньше некоторого критического значения х* просто не может существовать. Подобное соображение выглядит вполне реалистичным. Например, для очень мелких теплокровных животных, таких как колибри у птиц и землеройки у млекопитающих, дальнейшее уменьшение размеров создало бы серьезные проблемы с поддержанием температуры тела. По-видимому, колибри, питающиеся очень калорийным нектаром, уже достигли левой границы распределения птиц. Кстати, в покое они не в состоянии поддерживать постоянную температуру.
Пусть критическое значение признака соответствует интервалу x-8. Отсюда следует, что вид, занимающий этот самый левый интервал, может дать дочерний вид только с увеличенным размером особей, заселяя интервал х-7. С ходом времени мы получим распределение, которое будет весьма походить на предыдущее, отличаясь от него практически только отсутствием части, расположенной слева от «поглощающего экрана» (рис. 18, б). Естественно, с ходом времени среднее значение такого урезанного распределения будет смещаться вправо, а незначительным сдвигом его вершины (моды) можно пренебречь. Однако тщательный анализ реальных видовых распределений (подобных представленным на рис. 17) обнаруживает четкое смещение их вершины вправо, т. е. в сторону увеличения признака.
 Рис. 18. Моделирование динамики видовых распределений по логарифму величины количественного признака. а — вероятность изменения признака в обоих направлениях равна и не зависит от его величины; б — те же условия, но признак не может принимать значения меньше, чем x-8; в — ортоселекция (вероятность увеличения признака в 2 раза выше вероятности его уменьшения); г — рост адаптируемости (вероятность изменения признака в обоих направлениях равна и возрастает на 0,01 с переходом в соседний интервал справа); д — те же условия, что и в случае г (признак измеряется арифметической шкалой).
Рис. 18. Моделирование динамики видовых распределений по логарифму величины количественного признака. а — вероятность изменения признака в обоих направлениях равна и не зависит от его величины; б — те же условия, но признак не может принимать значения меньше, чем x-8; в — ортоселекция (вероятность увеличения признака в 2 раза выше вероятности его уменьшения); г — рост адаптируемости (вероятность изменения признака в обоих направлениях равна и возрастает на 0,01 с переходом в соседний интервал справа); д — те же условия, что и в случае г (признак измеряется арифметической шкалой).
Есть два пути получить динамику распределений, близкую к реальной. Первый состоит во введении асимметрии в процесс заселения смежных интервалов, второй — в ускорении заселения обоих соседних интервалов (вполне симметричного) по мере увеличения размеров особей. Первый путь — это ортоселекция, когда дочерний вид заселяет быстрее интервал справа, чем интервал слева. Если мы предположим, что вероятность заселения правого интервала в 2 раза выше, чем левого, то получим картину, представленную на рис. 18,в. Видно, что распределение быстро расползается вправо и в ту же сторону сдвигается его вершина. Обычно современные приверженцы синтетической теории эволюции считают, что за эти макроэволюционные тенденции ответственна именно ортоселекция. Их соображения выглядят примерно так. Увеличенные размеры тела дают индивидам очевидные преимущества: их бег становится быстрее и экономичнее, они лучше защищены от хищников и, наконец, они, обладая большей мышечной силой, имеют преимущества в драках с конкурентами за спаривание. Казалось бы все логично, хотя несколько антропоморфно. Действительно, людям свойственно воспевать крупных мужчин, их, так сказать, боевые качества. Попробуем посмотреть на это с иной точки зрения. Увеличенная масса тела затрудняет бег по пересеченной местности, к тому же крупный индивид требует для поддержания жизни больше пищи. Кроме того, с увеличением массы тела, безусловно, падает скорость реакции на внешние раздражители — весьма немаловажное обстоятельство для выживания. Едва ли существует положительная корреляция между размерами тела охотника и количеством приносимой им добычи — основы благополучия первобытного человека. Наконец, увеличенные размеры означают некоторое удлинение онтогенеза и соответственно снижение шансов достичь взрослой стадии. Таким образом, без точных оценок всех плюсов и минусов мы не в состоянии найти простую связь между размерами особи и ее приспособленностью. Единственной твердо установленной закономерностью, связывающей размеры тела животных с приспособленностью, является правило Бергмана, согласно которому среднепопуляционная масса тела у теплокровных животных одного вида увеличивается, с понижением среднегодовой температуры. Этому правилу подчиняется большинство видов птиц и млекопитающих, верно оно и для людей. Рассмотрим подробнее еще один путь, ведущий к результату, очень сходному с ожидаемым при ортоселекции. Пусть вероятность перехода дочерних видов в оба соседних интервала растет в прямой пропорции с номером интервала, в котором находился материнский вид. Заметим, что тем самым мы ввели предположение о существовании прямой зависимости между средним размером тела особи и адаптируемостью вида. На рис. 18, г представлена динамика таких распределений для случая, когда адаптируемость вида при переходе в соседний интервал справа возрастает на 0,01. Мы пришли к результату, фактически не отличимому от предыдущего: распределение расширяется вправо при переносе вправо и модального класса. Перейдя от логарифмической шкалы к арифметической (рис. 18, д), мы получим картину, весьма напоминающую динамику реальных распределений (см. рис. 17). Следовательно, наше исходное предположение о том, что вместе с увеличением средних размеров особей растет и адаптируемость вида, не противоречит фактам. Естественно, модель, которую мы здесь исследовали, весьма примитивна. Видовая дифференциация совсем не обязана всякий раз сочетаться с изменением размеров тела на какой-то фиксированный процент. Не вызывает сомнений, что часть видообразований происходит при изменении других признаков особей, и в этом случае виды-потомки остаются в классовых интервалах своих предков. Однако для правомочности рассмотренной модели достаточно, чтобы лишь некоторая доля всех актов видообразования сопровождалась изменением размеров тела. Конечно, еще легче сдвиг видовых распределений вправо объяснить с позиций ортогенеза (точнее, автогенеза), т. е. действием особых сил, детерминистических законов и стремлений, заложенных в самой природе организмов. В гл. 1 мы пытались вскрыть философско-психологические корни ортогенетических теорий. Приверженцы этих подкупающе простых идей даже вызывают симпатию своей верой в чудесные и таинственные силы природы. Однако ортогенез начисто лишен конструктивности и, по существу, несмотря на свою наукообразную форму, так же далек от объяснения эволюции, как астрология — от предсказания политических перемен. Итак, можно сформулировать главный вопрос нашего повествования: что же лежит в основе долговременных макроэволюционных тенденций — ортоселекция или рост адаптируемости? Для ответа попробуем перейти к генетической подоплеке этих явлений. Гены и мощность Попробуем ответить на вопрос, как изменится величина количественного признака при увеличении числа генов, управляющих его развитием. Всякая морфологическая структура состоит из клеток и внеклеточного вещества — продукта жизнедеятельности тех же клеток. Поэтому размеры структур, органов, да и всего организма определяются числом, объемом и экспортной активностью их клеток. Наличие гена мы определяем по его экспрессии. Если ген не активен ни в одной клетке организма, ни на одной стадии его развития, то мы можем считать, что его просто нет. Идеи о латентном (скрытом) существовании на протяжении миллионов лет вполне исправных генов ни на чем не основаны. Как селективно-нейтральные, латентные гены должны довольно быстро терять свое информационное содержание, поскольку не проверяются отбором. В чем же выражается генная активность? Прежде всего в транскрипции РНК, хотя это лишь необходимое, но далеко не достаточное условие для генной экспрессии. Обычно РНК должна «созреть» и транслироваться в полипептид, способный узнавать другие молекулы и, исполняя свою специфичную функцию, содействовать успеху организма в его борьбе за жизнь. Мы уже отмечали, что живое вещество можно рассматривать как катализатор, «пытающийся» ускорить собственное воспроизведение. При этом совокупное действие всех генов генома направлено на увеличение биомассы вида. Однако как быть с отдельными индивидами? Не станем же мы утверждать, что активность всех генов направлена на увеличение массы особи? Ответ здесь, казалось бы, должен быть отрицательным, но не станем спешить. Обычно особи многоклеточных организмов диплоидны, т. е. каждая хромосома их соматических клеток представлена двумя вариантами (гомологами), хотя для многих видов растений (да и некоторых животных) можно искусственным путем получить ряды форм с измененным числом гомологов. У гаплоидов это число равно одному, у триплоидов — трем, тетраплоидов — четырем и т. д. Оказалось, что триплоиды и тетраплоиды, как правило, крупнее (мощнее) диплоидов, а последние мощнее гаплоидов. Этот эффект обусловлен прежде всего соответствующим увеличением объема клеток всех типов. Таким образом, суммарный эффект генов направлен на прирост вещества цитоплазмы клеток. Еще в 1908 г. Р. Гертвиг установил «закон» ядерно-плазменного отношения, гласящий, что чем больше объем ядра, тем больше объем цитоплазмы. Но объем клеточного ядра в разных тканях одного организма определяется степенью деконденсации хроматина, которая прямо связана с активностью генов. Таким образом, этот «закон» также указывает на то, что суммарный эффект работы всех генов направлен на увеличение клеточной массы. Большинство мутаций с качественным эффектом на фенотип ведет к уменьшению размеров какой-либо морфологической структуры или всей массы тела особи. Примерно так же влияют и микроделеции, связанные с утратой гена. Увеличение размеров структур в результате серьезного повреждения гена наблюдается гораздо реже. У многих генетически изученных объектов известен ряд локусов, потеря или серьезное повреждение которых ведет к ступенчатой редукции одной и той же структуры. Например, у гороха известна большая серия подобных генов, определяющих массу семян. В качестве меры развития структуры следовало бы выбрать ее вклад в реализацию основных функций организма. Однако непонятно, как этот вклад измерить; гораздо проще в качестве искомой меры взять относительную величину структуры, т. е. отношение ее размера к размеру всего тела, олицетворяющего собой мощность всех функций организма. Как мы убедились выше (см. гл.2), сложность структуры коррелирует с ее мощностью. Многочисленные складки, карманы, выпячивания обеспечивают увеличение площади функционально активных поверхностей. Естественно, каждая складка появляется в определенном месте и в определенное время и является следствием «срабатывания» конкретного гена (или группы генов). Разнообразие клеточных элементов тоже коррелирует с генетической сложностью программы развития структуры. В итоге, возникает простая логическая цепь: чем больше генов контролирует развитие структуры, тем выше ее морфологическая сложность, чем выше эта сложность, тем больше ее относительные размеры и мощность. Все органы обладают рациональным строением, т. е. развивают максимальную мощность при минимальных энергетических затратах. Этот принцип симморфоза объясняет многократное возникновение в разных эволюционных стволах практически идентичных (в морфологическом отношении) структур, построенных как бы по одному плану. Поразительно, что тот же «план» довольно часто можно обнаружить и в конструкциях машин, созданных инженерной мыслью людей. Единственное разумное объяснение такой двойной конвергенции заключается в ограниченности спектра решений, обеспечивающих рабочую структуру максимальной мощностью при минимуме энергетических затрат. Рассмотрим такую техническую задачу, как идентификация объекта на расстоянии. Важность ее для организмов очевидна. Внешний предмет может быть и хищником, и жертвой, и особью противоположного пола, и совершенно нейтральным объектом. Если на него падает свет, то есть одно универсальное решение — зарегистрировать особенности отражения лучей от отдельных частей объекта. Очевидно, что для этого необходимо иметь светочувствительную структуру и устройство, направляющее на него отраженный свет. Всем этим снабжены многие оптические приборы, например фотокамера. Создавая такие приборы, человек руководствуется законами геометрической оптики и свойствами доступных ему материалов. В качестве светочувствительной структуры он использует фотоэмульсию, и фокусирует на нее лучи с помощью преломляющих свет прозрачных тел — прежде всего линз. Для получения четкого изображения объекта на фотоэмульсии мы должны уметь изменять или величину зазора между нею и линзой, или фокусное расстояние последней. Кроме того, мощность светового потока также нужно плавно регулировать, для чего оптические приборы снабжаются диафрагмой. Животные решают задачу идентификации удаленного предмета точно так же. Глаз с высокой разрешающей способностью обладает слоем светочувствительных клеток — сетчаткой — и всеми устройствами, которыми снабжен хороший оптический прибор: диафрагмой в виде радужины, системой линз (роговица, хрусталик, стекловидное тело), мышцами, способными изменять или фокусное расстояние линз, или их отстояние от сетчатки. Мощность органа зрения, таким образом, прямо зависит от светосилы системы его «линз», количества светочувствительных клеток сетчатки, эффективности устройств фокусирования лучей и регуляции интенсивности светового потока, а также от ряда других технических моментов. «Мощные», т. е. зоркие глаза устроены удивительно сходно у таких разных групп животных, как головоногие моллюски и высшие позвоночные. Орган зрения эти группы приобрели совершенно независимо, поскольку их эволюционные пути разошлись не позднее раннего кембрия от общего предка, который вместо глаз имел в лучшем случае лишь рассеянные по телу светочувствительные клетки. Соображения некоторых эволюционистов о возникновении глаз под действием неких внутренних факторов, толкающих орган зрения по пути прогресса, в настоящее время выглядят по меньшей мере анахронизмом. Специальные гены в эволюции Адаптируемость — это способность вида поднять уровень своей абсолютной приспособленности в неоптимальных условиях среды. Что же такое абсолютная приспособленность вида и как определить ее уровень? Пока не существует строгого определения этих понятий. По-видимому, лучше всего представление об абсолютной приспособленности передает величина биомассы (или количество особей), производимой видом за единицу времени. Очевидно, что эта характеристика прямо связана с биомассой вида (или с численностью его особей). Для того чтобы проследить за динамикой уровня абсолютной приспособленности вида, мы должны иметь возможность наблюдать за ним в течение многих десятков (и даже сотен) поколений. Фактически нам следовало бы наблюдать всего за двумя характеристиками вида — географическим ареалом и числом особей в его популяциях. Обычно плотность популяций максимальна в центре ареала, откуда некоторый «избыток» особей перебрасывается миграционным потоком на периферию, где гибнет под влиянием неблагоприятных факторов. Если мощность производства биомассы в центре ареала ослабевает, то падает средняя плотность популяций и сокращается площадь видового ареала. Конечно, при ежегодных наблюдениях мы бы заметили довольно резкие колебания обеих характеристик, но если бы их средние многолетние значения достоверно не снижались, то можно было бы сказать, что данный вид неплохо приспособлен к среде своего обитания. Теперь проведем один мысленный эксперимент: истребим всех особей вида, кроме одной оплодотворенной самки. Поместим ее в искусственно созданные идеальные условия, многократно размножим потомков и выпустим их обратно в места обитания предков. Едва ли кто усомнится в исходе этого предприятия: вероятнее всего, вид вскоре восстановит свой прежний ареал и численность. В успехе подобного эксперимента нас убеждают многочисленные случаи восстановления совсем было истребленных видов (выхухоль, бобр, зубр, бизон и т. д.). Кроме того, о силе жизни, заключенной в считанных штуках особей, свидетельствуют примеры успешной интродукции вида (иногда совсем нежелательной) в новые места обитания. Кролики в Австралии, лошади в Америке, колорадский жук в Европе, рапана в Черном море и масса других подобных случаев. Везде мы видим, что основные характеристики, определяющие абсолютную приспособленность вида, заключены в геноме типичной, т. е. фактически любой особи. Пожалуй, наиболее ярко об этом свидетельствует «успех» канадской элодеи, впервые обнаруженной в Ирландии в 1836 г. У себя на родине эта элодея представлена мужскими и женскими растениями, т. е. ей вполне свойственно половое размножение, кроме того, каждое растение (точнее, любой фрагмент его стебля с несколькими мутовками) способно к неограниченному росту и ветвлению. Завезенный в Европу женский экземпляр, размножаясь только вегетативно, породил столько потомков, что их биомасса вскоре стала мешать судоходству в каналах Англии. В настоящее время канадская элодея уже достигла рек и озер Восточной Сибири. Сила этого единственного генотипа отражена в названии, данному растению в Европе, — «водяная чума». Правда, сейчас его мощь явно ослабела, вероятно, стал сказываться мутационный груз, для сброса которого нужен половой процесс. По-видимому, главное, что несет в себе типичная особь вида, заключается в скорости, с которой организм в конкретных условиях среды воспринимает извне вещество и энергию, преобразуя их в биомассу потомков. Что же определяет эту скорость? Очевидно, только мощность рабочих структур типичной особи. Напомним, что в гл. 2 мы определили такую мощность, как объем работы, выполняемой структурой за единицу времени в режиме максимальной нагрузки. При ухудшении среды мощность какой-то рабочей структуры может оказаться недостаточной, поэтому численность вида и его ареал сократятся, и восстановить их будет возможно, лишь усилив мощность лимитирующей структуры, ставшей фактически одной из главных компонент приспособленности. Выходит, абсолютную приспособленность вида можно было бы выразить в единицах мощности. В данной ситуации мутации, повышающие мощность лимитирующей структуры, очевидно становятся благоприятными, так как они повышают уровень относительной приспособленности их носителей. Поэтому коэффициент отбора (s) в пользу этих мутаций должен быть близок к относительному приросту мощности лимитирующей структуры, во всяком случае, он должен быть ему пропорционален, т. е.
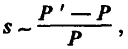 (5.3)
где Р' — мощность структуры у особей — носителей благоприятной мутации; Р — мощность той же структуры у средней особи популяции. Фактически это означает, что фиксация каждой благоприятной мутации увеличивает мощность лимитирующей структуры в некоторое число раз (на какой-то процент). При продолжающемся ухудшении среды продолжается и процесс накопления благоприятных мутаций. В итоге мощность лимитирующей структуры нарастает вдоль филетического маршрута как ряд:
(5.3)
где Р' — мощность структуры у особей — носителей благоприятной мутации; Р — мощность той же структуры у средней особи популяции. Фактически это означает, что фиксация каждой благоприятной мутации увеличивает мощность лимитирующей структуры в некоторое число раз (на какой-то процент). При продолжающемся ухудшении среды продолжается и процесс накопления благоприятных мутаций. В итоге мощность лимитирующей структуры нарастает вдоль филетического маршрута как ряд:
 (5.4)
где Р0 — мощность структуры до начала действия движущего отбора; xi — число раз, в которое увеличивается мощность структуры в ходе фиксации i-й мутации. Это как раз те «незначительные изменения», аккумуляция которых, по выражению Дарвина, ведет к созданию органа «любой мыслимой степени совершенства».
Заметим, что логарифм мощности структуры (lnP) после фиксации и благоприятных мутаций определяется равенством
(5.4)
где Р0 — мощность структуры до начала действия движущего отбора; xi — число раз, в которое увеличивается мощность структуры в ходе фиксации i-й мутации. Это как раз те «незначительные изменения», аккумуляция которых, по выражению Дарвина, ведет к созданию органа «любой мыслимой степени совершенства».
Заметим, что логарифм мощности структуры (lnP) после фиксации и благоприятных мутаций определяется равенством
 (5.5)
где стоящий под знаком суммы lnxi представляет собой вклад i-й благоприятной мутации в логарифм мощности рабочей структуры; для удобства дальнейшего изложения этот вклад будем обозначать аi'. Если единицей измерения мощности рабочей структуры выбрать ее исходную величину Р0, то (учитывая, что lnl = 0) уравнение (5.5) можно представить в более простой форме:
(5.5)
где стоящий под знаком суммы lnxi представляет собой вклад i-й благоприятной мутации в логарифм мощности рабочей структуры; для удобства дальнейшего изложения этот вклад будем обозначать аi'. Если единицей измерения мощности рабочей структуры выбрать ее исходную величину Р0, то (учитывая, что lnl = 0) уравнение (5.5) можно представить в более простой форме:
 (5.6)
Напомним, что аi' — вклад i-й мутации, зафиксированной отбором, направленным на усиление лимитирующей структуры.
Теперь введем одно определение: локусы, по которым шло замещение новых аллелей, повышающих мощность структуры в ее историческом развитии, назовем специальными генами структуры. Вполне очевидно, что некоторые из этих генов могут повторно мутировать, порождая новые аллели, усиливающие лимитирующую функцию. Таким образом, среди n благоприятных мутаций, зафиксированных движущим отбором, далеко не все связаны с увеличением числа специальных генов N.
Представим себе, что изменение среды приняло противоположное направление. В новых условиях мощность структуры, развитой за предыдущий период, может стать избыточной, поэтому ничто не будет препятствовать накоплению мутаций, ослабляющих экспрессию ее специальных генов. Более того, энергия и вещество, которые ранее расходовались на поддержание высокой мощности этой структуры, теперь могут быть направлены на создание дополнительной биомассы, т. е. на повышение абсолютной приспособленности вида. Таким образом, ослабление рабочей структуры в данном случае представляет собой адаптивный ответ вида на «требование» изменившейся среды. Через некоторое время новая среда может снова потребовать усиления функции той же структуры, и снова благоприятными станут мутации, усиливающие экспрессию ее специальных генов.
Таким образом, для вида, предки которого прошли через многократные изменения требований среды к интенсивности функции определенной рабочей структуры, логарифм ее мощности можно представить в виде суммы вкладов N специальных генов, где вклад каждого такого гена (его экспрессия) (а) может изменяться от нуля до какого-то максимального значения, т. е.
(5.6)
Напомним, что аi' — вклад i-й мутации, зафиксированной отбором, направленным на усиление лимитирующей структуры.
Теперь введем одно определение: локусы, по которым шло замещение новых аллелей, повышающих мощность структуры в ее историческом развитии, назовем специальными генами структуры. Вполне очевидно, что некоторые из этих генов могут повторно мутировать, порождая новые аллели, усиливающие лимитирующую функцию. Таким образом, среди n благоприятных мутаций, зафиксированных движущим отбором, далеко не все связаны с увеличением числа специальных генов N.
Представим себе, что изменение среды приняло противоположное направление. В новых условиях мощность структуры, развитой за предыдущий период, может стать избыточной, поэтому ничто не будет препятствовать накоплению мутаций, ослабляющих экспрессию ее специальных генов. Более того, энергия и вещество, которые ранее расходовались на поддержание высокой мощности этой структуры, теперь могут быть направлены на создание дополнительной биомассы, т. е. на повышение абсолютной приспособленности вида. Таким образом, ослабление рабочей структуры в данном случае представляет собой адаптивный ответ вида на «требование» изменившейся среды. Через некоторое время новая среда может снова потребовать усиления функции той же структуры, и снова благоприятными станут мутации, усиливающие экспрессию ее специальных генов.
Таким образом, для вида, предки которого прошли через многократные изменения требований среды к интенсивности функции определенной рабочей структуры, логарифм ее мощности можно представить в виде суммы вкладов N специальных генов, где вклад каждого такого гена (его экспрессия) (а) может изменяться от нуля до какого-то максимального значения, т. е.
 (5.7)
Это выражение удобнее представить в несколько иной форме:
lnP = Na, (5.8)
где а — уровень средней экспрессии специального гена, т. е. его средний вклад в логарифм мощности рабочей структуры. У разных видов а может варьировать в очень широком диапазоне: от нуля, когда все специальные гены «выключены», до величины, соответствующей уровню их максимальной активности. Ясно, что уровень средней экспрессии специального гена рабочей структуры зависит от предыстории вида, от того, как долго среда «требовала» от предков усиления мощности структуры и как долго была к ней безразлична или даже «ждала» ослабления.
Пусть мы имеем дело с совокупностью родственных видов одной филетической группы. Рассмотрим у них какую-нибудь гомологичную структуру с одной и той же функцией. У каждого вида (в соответствии с (5.8)) логарифм мощности структуры равен произведению Na. Теперь перейдем к среднему (для видов филетической группы) логарифму мощности структуры (<lnP>), т. е. от каждого вида возьмем значение логарифма мощности структуры ее типичной особи, полученные величины сложим, а их сумму разделим на число видов в группе. В теории вероятностей доказывается, что среднее значение произведения независимо варьирующих сомножителей равно произведению их средних значений. Следовательно, средневидовой логарифм мощности структуры будет равен средневидовому уровню экспрессии ее специальных генов (<а>), умноженному на среднее число этих генов у одного вида (<N>), т. е.
<lnP> = <N> <а>. (5.9)
Если линии филетической группы уже неоднократно сталкивались с изменениями требований среды к мощности какой-то рабочей структуры, то средневидовой уровень экспрессии ее специальных генов можно считать в какой-то степени стабилизированным. В этом случае соотношение (5.9) упрощается до
<lnP> ~ <N>. (5.10)
Число специальных генов, определяющих развитие структуры, можно принять за меру ее генетической сложности, поэтому соотношение (5.10) означает, что средневидовая мощность структуры, измеренная логарифмической шкалой, пропорциональна ее средневидовой генетической сложности.
Специальные гены в онтогенезе
Нетрудно убедиться в том, что практически каждый элемент сложной рабочей структуры служит усилению ее мощности. Возьмем, к примеру, сердце млекопитающих. Здесь мы видим четыре отдела, четко разделенных по своей функции, несколько весьма сложно устроенных клапанов, особый нервно-мышечный аппарат для обеспечения ритмических сокращений, да и сама ткань сердечной мышцы поражает необычностью своего строения. Повреждение любой из перечисленных структур ведет к ослаблению сердца как насоса и сопровождается симптомами сердечной недостаточности.Той же цели усиления мощности служат и все рассмотренные выше устройства глаза, повышающие остроту зрения.
Еще в гл. 2 мы пришли к заключению, что увеличение массы тела животного ведет к усложнению строения органов, особенно если их мощность зависит от каких-нибудь одно- или двухмерных структур. Любой орган возникает в ходе индивидуального развития, проходя последовательно ряд стадий, — от исходного зачатка до дефинитивного (взрослого) состояния. Из генетики развития хорошо известно, что может быть найдена мутация, прерывающая этот процесс на любой его стадии. Вполне очевидно, что чем больше качественно различных стадий проходит зачаток в онтогенезе, тем больше генов должна включать в себя программа его развития и тем сложнее, а следовательно, и мощнее будет дефинитивная структура. Если гены, управляющие развитием структуры, назвать специальными, то фактически мы снова получаем тот же результат: при возрастании числа специальных генов увеличивается мощность рабочей структуры.
Качественная эквивалентность обоих подходов покоится на так называемом «основном биогенетическом законе» Геккеля — Мюллера, суть которого передается броской формулировкой Э. Геккеля: «Онтогенез всякого организма есть краткое и сжатое повторение филогенеза данного вида». Такое повторение напоминает как бы перечень основных «глав» филогенеза, что объясняет другое название этого явления — рекапитуляция (от латинского capitulum — глава книги).
В формулировке основного биогенетического закона есть нечто мистическое, прямо-таки роковое, к тому же он указывает на подкупающе простой способ восстановления исторического развития организмов, как будто для этого достаточно лишь изучить вполне доступный наблюдению онтогенез. Однако было бы крайне наивно искать в раннем эмбриогенезе человека стадии рептилий, амфибий и рыб, а в нашем младенчестве и детстве — стадии низших обезьян, питекантропа и неандертальца. Тем не менее явление рекапитуляции существует и нуждается в объяснении.
По-видимому, оно основано на том, что филогенез «творится» в ходе онтогенеза. При этом всякая новизна, приведшая к увеличению мощности дефинитивной структуры, обусловлена вполне определенным изменением течения онтогенеза. Довольно часто оно состоит в довершении процесса развития еще одной стадией. Такой путь внедрения эволюционных инноваций А.Н. Северцов назвал анаболиями (т. е. надставками). Несколько реже изменения касаются средних и начальных стадий развития, которые соответствуют северцовским «девиациям» и «архалаксисам».
Ранее мы уже отмечали, что мутации, затрагивающие поздние стадии развития структур, должны фиксироваться чаще, поскольку они в меньшей степени влияют на развитие других структур, т. е., как правило, уровень «благоприятности» анаболий должен быть выше, чем девиаций и тем более архалаксисов. Заметим, что основной биогенетический закон не знал бы исключений, если бы эволюционная новизна всегда возникала только в виде надставок к конечной стадии программы развития предка. Поскольку каждая надставка ведет к усилению рабочей структуры, то фактически это тот же процесс, который мы рассмотрели в предыдущем параграфе (см. (5.4)). Только там речь шла о фиксации отдельных благоприятных мутаций с очень небольшим эффектом, тогда как каждая анаболия — это, скорее всего, серия подобных фиксаций.
Пока мы не получили никаких неожиданных результатов. Все выглядит вполне тривиально, и остается совершенно непонятным, какая сила заставляет увеличивать для некоторых рабочих структур число специальных генов при долговременных эволюционных тенденциях.
Лопастная линия аммоноидей
Уже более 100 лет внимание эволюционистов приковано к одной замечательной тенденции, сопровождающей эволюцию аммоноидей, — усложнению их лопастной линии (см. рис. 3). Эта филетическая группа возникла в раннем девоне, достигла огромного таксономического разнообразия в мезозое и полностью вымерла одновременно с динозаврами в конце мелового периода, просуществовав на Земле около 330 миллионов лет.
Аммоноидеи относятся к головоногим моллюскам с наружной раковиной — конической известковой трубкой, свернутой в логарифмическую спираль (рис. 19). Тело моллюска находилось в широкой части трубки — так называемой жилой камере, остальная, нежилая, часть раковины (фрагмакон) была разделена на герметичные отсеки многочисленными поперечными перегородками из перламутра. Отсеки заполнялись газом с давлением, близким к атмосферному, поэтому при погружении на большую глубину наружные стенки раковины испытывали изрядное давление воды, стремящееся раздавить пустотелую часть раковины. Газ в отсеках был необходим для уравновешивания животного в воде. Точная балансировка обеспечивалась специальным жгутообразным органом — сифоном, пронизывающим все поперечные перегородки.
Поперечные перегородки почти наверняка играли роль корабельных шпангоутов, т. е. выполняли функцию крепежных конструкций, работающих на сжатие. Напряжение наружной стенки передавалось на поперечную перегородку через линию их срастания. Эта знаменитая линия, которая очерчивает контур периферического края перегородки, носит разные названия: сутура, перегородочная, лопастная, шовная.
Представим себе, что условия среды «вынуждают» вид переселиться на большую глубину, где на каждый элемент поверхности раковины будет действовать повышенное гидростатическое давление. Ясно, что для успеха такого переселения нужно позаботиться об укреплении раковины. Этого можно достичь четырьмя способами: 1) утолщить стенку раковины; 2) увеличить число поперечных перегородок; 3) уменьшить размеры раковины (так как критическое напряжение, разрушающее полый шар, обратно пропорционально его радиусу); 4) изогнуть лопастную линию, что приведет к ее удлинению без изменения размеров раковины.
Первые два пути ведут к увеличению инерционной массы животного, что должно отрицательно сказаться на маневренности его плавания и, кроме того, утяжеленную особь труднее уравновесить газом в пустых отсеках. Третье средство вполне приемлемо, но тогда животному надо проститься со всеми преимуществами крупных размеров. А что делать, если новая более глубинная среда требует к тому же и увеличения массы тела? Тогда остается только четвертый путь. Действительно, удлиненная за счет изгибов лопастная линия может принять напряжение с большей поверхности, при этом сама раковина не утяжеляется. Именно это решение выглядит наиболее заманчиво, когда среда «требует» увеличения размеров индивида. При этом для компенсации возрастающего давления воды рост длины лопастной линии должен быть аллометричным, в идеале он должен следовать за квадратом радиуса раковины. Во всяком случае, при укрупнении особей должна возрастать степень изогнутости сутуры, ее сложность.
(5.7)
Это выражение удобнее представить в несколько иной форме:
lnP = Na, (5.8)
где а — уровень средней экспрессии специального гена, т. е. его средний вклад в логарифм мощности рабочей структуры. У разных видов а может варьировать в очень широком диапазоне: от нуля, когда все специальные гены «выключены», до величины, соответствующей уровню их максимальной активности. Ясно, что уровень средней экспрессии специального гена рабочей структуры зависит от предыстории вида, от того, как долго среда «требовала» от предков усиления мощности структуры и как долго была к ней безразлична или даже «ждала» ослабления.
Пусть мы имеем дело с совокупностью родственных видов одной филетической группы. Рассмотрим у них какую-нибудь гомологичную структуру с одной и той же функцией. У каждого вида (в соответствии с (5.8)) логарифм мощности структуры равен произведению Na. Теперь перейдем к среднему (для видов филетической группы) логарифму мощности структуры (<lnP>), т. е. от каждого вида возьмем значение логарифма мощности структуры ее типичной особи, полученные величины сложим, а их сумму разделим на число видов в группе. В теории вероятностей доказывается, что среднее значение произведения независимо варьирующих сомножителей равно произведению их средних значений. Следовательно, средневидовой логарифм мощности структуры будет равен средневидовому уровню экспрессии ее специальных генов (<а>), умноженному на среднее число этих генов у одного вида (<N>), т. е.
<lnP> = <N> <а>. (5.9)
Если линии филетической группы уже неоднократно сталкивались с изменениями требований среды к мощности какой-то рабочей структуры, то средневидовой уровень экспрессии ее специальных генов можно считать в какой-то степени стабилизированным. В этом случае соотношение (5.9) упрощается до
<lnP> ~ <N>. (5.10)
Число специальных генов, определяющих развитие структуры, можно принять за меру ее генетической сложности, поэтому соотношение (5.10) означает, что средневидовая мощность структуры, измеренная логарифмической шкалой, пропорциональна ее средневидовой генетической сложности.
Специальные гены в онтогенезе
Нетрудно убедиться в том, что практически каждый элемент сложной рабочей структуры служит усилению ее мощности. Возьмем, к примеру, сердце млекопитающих. Здесь мы видим четыре отдела, четко разделенных по своей функции, несколько весьма сложно устроенных клапанов, особый нервно-мышечный аппарат для обеспечения ритмических сокращений, да и сама ткань сердечной мышцы поражает необычностью своего строения. Повреждение любой из перечисленных структур ведет к ослаблению сердца как насоса и сопровождается симптомами сердечной недостаточности.Той же цели усиления мощности служат и все рассмотренные выше устройства глаза, повышающие остроту зрения.
Еще в гл. 2 мы пришли к заключению, что увеличение массы тела животного ведет к усложнению строения органов, особенно если их мощность зависит от каких-нибудь одно- или двухмерных структур. Любой орган возникает в ходе индивидуального развития, проходя последовательно ряд стадий, — от исходного зачатка до дефинитивного (взрослого) состояния. Из генетики развития хорошо известно, что может быть найдена мутация, прерывающая этот процесс на любой его стадии. Вполне очевидно, что чем больше качественно различных стадий проходит зачаток в онтогенезе, тем больше генов должна включать в себя программа его развития и тем сложнее, а следовательно, и мощнее будет дефинитивная структура. Если гены, управляющие развитием структуры, назвать специальными, то фактически мы снова получаем тот же результат: при возрастании числа специальных генов увеличивается мощность рабочей структуры.
Качественная эквивалентность обоих подходов покоится на так называемом «основном биогенетическом законе» Геккеля — Мюллера, суть которого передается броской формулировкой Э. Геккеля: «Онтогенез всякого организма есть краткое и сжатое повторение филогенеза данного вида». Такое повторение напоминает как бы перечень основных «глав» филогенеза, что объясняет другое название этого явления — рекапитуляция (от латинского capitulum — глава книги).
В формулировке основного биогенетического закона есть нечто мистическое, прямо-таки роковое, к тому же он указывает на подкупающе простой способ восстановления исторического развития организмов, как будто для этого достаточно лишь изучить вполне доступный наблюдению онтогенез. Однако было бы крайне наивно искать в раннем эмбриогенезе человека стадии рептилий, амфибий и рыб, а в нашем младенчестве и детстве — стадии низших обезьян, питекантропа и неандертальца. Тем не менее явление рекапитуляции существует и нуждается в объяснении.
По-видимому, оно основано на том, что филогенез «творится» в ходе онтогенеза. При этом всякая новизна, приведшая к увеличению мощности дефинитивной структуры, обусловлена вполне определенным изменением течения онтогенеза. Довольно часто оно состоит в довершении процесса развития еще одной стадией. Такой путь внедрения эволюционных инноваций А.Н. Северцов назвал анаболиями (т. е. надставками). Несколько реже изменения касаются средних и начальных стадий развития, которые соответствуют северцовским «девиациям» и «архалаксисам».
Ранее мы уже отмечали, что мутации, затрагивающие поздние стадии развития структур, должны фиксироваться чаще, поскольку они в меньшей степени влияют на развитие других структур, т. е., как правило, уровень «благоприятности» анаболий должен быть выше, чем девиаций и тем более архалаксисов. Заметим, что основной биогенетический закон не знал бы исключений, если бы эволюционная новизна всегда возникала только в виде надставок к конечной стадии программы развития предка. Поскольку каждая надставка ведет к усилению рабочей структуры, то фактически это тот же процесс, который мы рассмотрели в предыдущем параграфе (см. (5.4)). Только там речь шла о фиксации отдельных благоприятных мутаций с очень небольшим эффектом, тогда как каждая анаболия — это, скорее всего, серия подобных фиксаций.
Пока мы не получили никаких неожиданных результатов. Все выглядит вполне тривиально, и остается совершенно непонятным, какая сила заставляет увеличивать для некоторых рабочих структур число специальных генов при долговременных эволюционных тенденциях.
Лопастная линия аммоноидей
Уже более 100 лет внимание эволюционистов приковано к одной замечательной тенденции, сопровождающей эволюцию аммоноидей, — усложнению их лопастной линии (см. рис. 3). Эта филетическая группа возникла в раннем девоне, достигла огромного таксономического разнообразия в мезозое и полностью вымерла одновременно с динозаврами в конце мелового периода, просуществовав на Земле около 330 миллионов лет.
Аммоноидеи относятся к головоногим моллюскам с наружной раковиной — конической известковой трубкой, свернутой в логарифмическую спираль (рис. 19). Тело моллюска находилось в широкой части трубки — так называемой жилой камере, остальная, нежилая, часть раковины (фрагмакон) была разделена на герметичные отсеки многочисленными поперечными перегородками из перламутра. Отсеки заполнялись газом с давлением, близким к атмосферному, поэтому при погружении на большую глубину наружные стенки раковины испытывали изрядное давление воды, стремящееся раздавить пустотелую часть раковины. Газ в отсеках был необходим для уравновешивания животного в воде. Точная балансировка обеспечивалась специальным жгутообразным органом — сифоном, пронизывающим все поперечные перегородки.
Поперечные перегородки почти наверняка играли роль корабельных шпангоутов, т. е. выполняли функцию крепежных конструкций, работающих на сжатие. Напряжение наружной стенки передавалось на поперечную перегородку через линию их срастания. Эта знаменитая линия, которая очерчивает контур периферического края перегородки, носит разные названия: сутура, перегородочная, лопастная, шовная.
Представим себе, что условия среды «вынуждают» вид переселиться на большую глубину, где на каждый элемент поверхности раковины будет действовать повышенное гидростатическое давление. Ясно, что для успеха такого переселения нужно позаботиться об укреплении раковины. Этого можно достичь четырьмя способами: 1) утолщить стенку раковины; 2) увеличить число поперечных перегородок; 3) уменьшить размеры раковины (так как критическое напряжение, разрушающее полый шар, обратно пропорционально его радиусу); 4) изогнуть лопастную линию, что приведет к ее удлинению без изменения размеров раковины.
Первые два пути ведут к увеличению инерционной массы животного, что должно отрицательно сказаться на маневренности его плавания и, кроме того, утяжеленную особь труднее уравновесить газом в пустых отсеках. Третье средство вполне приемлемо, но тогда животному надо проститься со всеми преимуществами крупных размеров. А что делать, если новая более глубинная среда требует к тому же и увеличения массы тела? Тогда остается только четвертый путь. Действительно, удлиненная за счет изгибов лопастная линия может принять напряжение с большей поверхности, при этом сама раковина не утяжеляется. Именно это решение выглядит наиболее заманчиво, когда среда «требует» увеличения размеров индивида. При этом для компенсации возрастающего давления воды рост длины лопастной линии должен быть аллометричным, в идеале он должен следовать за квадратом радиуса раковины. Во всяком случае, при укрупнении особей должна возрастать степень изогнутости сутуры, ее сложность.
 Рис. 19. Раковина аммонита. а — общий вид; б — изолированный фрагмокон. Видны лопастные линии и поверхность перегородки, отделяющей жилую камеру от фрагмокона.
Рис. 19. Раковина аммонита. а — общий вид; б — изолированный фрагмокон. Видны лопастные линии и поверхность перегородки, отделяющей жилую камеру от фрагмокона.
Заметим, что та же проблема вставала и при росте каждой особи. Аммоноидеи начинали свой онтогенез с очень маленького яйца диаметром меньше одного миллиметра, из которого вылуплялась крошечная личинка — амонителла, уже обладавшая раковинкой с двумя перегородками. По мере роста животному приходилось все время надстраивать и расширять свою жилую камеру. При этом сам моллюск передвигался ближе к устью и сооружал за собой новую поперечную перегородку. Таким образом, каждая раковина взрослого животного содержала весь набор перегородок, построенных им за свою жизнь. Это обстоятельство позволяет проследить за изменением лопастной линии в ходе индивидуального развития. Любая последовательность этих линий, упорядоченная по возрасту (онтограмма), демонстрирует их усложнение в онтогенезе (рис. 20). Еще раз заметим, что перегородка сделана из очень прочного, совсем не пластичного материала, и изогнуть ее периферический край чисто механически невозможно. Элементами сложности сутуры являются ее изгибы. Одни из них, обращенные вогнутостью к устью раковины, называют лопастями, другие, изогнутые в противоположную сторону, — седлами. На основании анализа множества вариантов усложнения сутуры в ходе онтогенеза создается впечатление, что каждый изгиб лопастной линии возникал на месте вполне определенного элемента более молодой особи. Углублению изгиба обычно предшествовала фаза рассечения сутуры (ее излома) в совершенно конкретном месте вполне определенного элемента. Известковое вещество перегородки выделялось специальными клетками мантии моллюска, поэтому за каждым элементом сутуры наверняка стояла особым образом дифференцированная область мантии. Следовательно, появление каждого нового элемента — это новый шаг в морфологической дифференциации мантии. Сопоставляя онтограммы аммоноидей из одних и тех же филетических пучков на разных этапах их исторического развития, можно заметить, что пути развития изгибов лопастной линии в онтогенезе в основных чертах повторяют пути их филетических предков. При этом увеличение сложности в онтогенезе потомков, как правило, шло за счет анаболий, т. е. добавления новых элементов к взрослой сутуре предков. Таким образом, онтограммы лопастных линий аммоноидей могут служить прекрасной иллюстрацией к явлению рекапитуляции. Вместе с тем, наверняка всегда сохранялось требование отбора к сокращению длительности индивидуального развития, поэтому более «прогрессивные» формы примерно за одно и то же время онтогенеза должны были построить более сложную лопастную линию. Это объясняет тенденцию к переносу взрослых стадий предков на все более ранние обороты раковины потомков. В конце концов зачатки некоторых элементов сутуры взрослых девонских аммоноидей можно обнаружить уже на лопастной линии мезозойской амонителлы.
 Рис. 20. Усложнение лопастной линии в онтогенезе мелового аммонита Gaudriceras tenuiliratum (по: [Михайлова, 1983]).
а — д — последовательные стадии развития.
Рис. 20. Усложнение лопастной линии в онтогенезе мелового аммонита Gaudriceras tenuiliratum (по: [Михайлова, 1983]).
а — д — последовательные стадии развития.
У палеозойских аммоноидей к рассечению были способны только лопасти, причем существовало несколько вариантов их усложнения. Рассечение седел впервые возникло в мезозое и сначала оно затронуло лишь их склоны. Позже всего аммоноидеи «освоили» рассечение вершин седел. Качественный анализ усложнения сутуры в ходе эволюции аммоноидей показывает, что вплоть до начала юры во всех крупных филогенетических стволах шло увеличение числа способов усложнения сутуры. Обычно эволюционно новый способ появлялся в конце онтогенеза и затем начинал «срабатывать» на более ранних стадиях индивидуального развития филетических потомков. Следует отметить, что однажды возникший способ усложнения и перенесенный затем в глубь онтогенеза потомков становился своеобразным маркером филетического пучка. Не вызывает сомнения, что за каждым способом усложнения сутуры стояли определенные гены, управлявшие развитием железистых клеток различных участков мантии. Таким образом, качественный анализ явления показывает, что в ходе исторического развития аммоноидей действовал процесс аккумуляции генов, осуществлявших усложнение лопастной линии, их можно назвать специальными генами сутуры. Усложнение лопастной линии В качестве меры изогнутости лопастной линии можно выбрать отношение l/l0, где l — длина сутуры, а l0 — длина контура поперечного среза раковинной трубки. Длину l0 имела бы сутура, если реальную перегородку с ее сложным рельефом заменить на совершенно плоскую пластинку. Аммоноидеи произошли от бактритоидей — головоногих моллюсков с прямой толстостенной раковиной. Их поперечные перегородки были практически плоскими и имели ровный периферический край, т. е. для них l~l0. Вспоминая наше определение мощности (см. гл.2), можно считать степень изогнутости сутуры (l/l0) мерой ее мощности (как способности противостоять гидростатическому давлению на стенку раковины). Поскольку естественный отбор «измеряет» величину рабочих структур как бы логарифмической линейкой, в качестве меры сложности лопастной линии Z имеет смысл выбрать величину логарифма ее мощности, т. е. Z = ln(l/l0). Заметим, что у бактритоидей сложность лопастной линии близка к нулю.
 Рис. 21. Динамика видовых распределений аммоноидей по сложности лопастной линии в ходе исторического развития.
Рис. 21. Динамика видовых распределений аммоноидей по сложности лопастной линии в ходе исторического развития.
Рассмотрим, как изменялись в процессе эволюции аммоноидей видовые распределения по сложности лопастной линии (рис. 21). Первое, что бросается в глаза, это увеличение их размаха. За 300 млн лет от раннего девона до позднего мела размах распределений возрос в 13 раз! При этом правый их край продвинулся вправо гораздо дальше левого. Форма распределений изменялась не слишком радикально, хотя в целом юрские и меловые распределения симметричнее палеозойских и триасских, для которых характерен некоторый избыток видов с упрощенной сутурой. Создается впечатление, что всю картину усложнения лопастной линии можно свести к растяжению исходных распределений вдоль горизонтальной полуоси. Заметим, что при таком растяжении среднее значение и сигма должны возрастать согласованным образом (рис. 22, а). Видно, что в течение четырех геологических периодов сутура неуклонно усложнялась, однако за юрский и меловой периоды серьезного подъема средней сложности (<Z>) не произошло. Степень согласованности изменений среднего значения и сигмы выражается в очень высоком значении коэффициента корреляции (рис. 23), фактически зависимость этих параметров друг от друга близка к линейной. Заметим, что среднее значение сложности лопастной линии (<Z>) должно быть пропорциональным среднему числу специальных генов этой структуры, накопленных филетической группой за время ее эволюции от бактритоидей (см. (5.10)), т. е. <Z> ~ <N>.
 Рис. 22. Изменение средней сложности (<Z>) и межвидовой изменчивости по сложности сутуры (σz) аммоноидей в их историческом развитии, а — кривые роста <Z> и σz для всех аммоноидей; б — рост средней сложности лопастной линии в четырех филетических группах аммоноидей. 1 — агониатиты, 2 — гониатиты, 3 — цератиты, 4 — аммониты.
D — девон, С — карбон, Р — пермь, Т — триас, J — юра, К — мел.
Рис. 22. Изменение средней сложности (<Z>) и межвидовой изменчивости по сложности сутуры (σz) аммоноидей в их историческом развитии, а — кривые роста <Z> и σz для всех аммоноидей; б — рост средней сложности лопастной линии в четырех филетических группах аммоноидей. 1 — агониатиты, 2 — гониатиты, 3 — цератиты, 4 — аммониты.
D — девон, С — карбон, Р — пермь, Т — триас, J — юра, К — мел.
Таким образом, график изменения средней сложности лопастной линии показывает, что вплоть до начала юры шло неуклонное накопление специальных генов и оно прекратилось вместе с прекращением увеличения числа способов рассечения сутуры. При этом подъем <Z> наблюдался во всех четырех крупных филетических группах аммоноидей: агониатитов, гониатитов, цератитов и аммонитов (см. рис. 22,б). Изломы на кривой роста средней сложности сутуры аммоноидей (см. рис. 22) отражают моменты вымираний отдельных филетических групп. Например, пермско-триасская катастрофа покончила с гониатитами и большинством агониатитов, когда у обеих групп средняя сложность сутуры достигла очень высокого уровня развития. Лопастная линия выживших цератитов оставалась еще очень простой.
 Рис. 23. Связь между средним значением (<Z>) и сигмой (σ) для распределений аммоноидей по сложности лопастной линии.
1 — девон; 2 — карбон; 3 — пермь; 4 — триас; 5 — юра; 6 — мел.
Рис. 23. Связь между средним значением (<Z>) и сигмой (σ) для распределений аммоноидей по сложности лопастной линии.
1 — девон; 2 — карбон; 3 — пермь; 4 — триас; 5 — юра; 6 — мел.
Конечно, сам факт подъема степени изогнутости лопастной линии в течение примерно 200 млн лет весьма впечатляет и является великолепной иллюстрацией к эволюционному прогрессу. В то же время ничуть не менее поражает застой в усложнении этой структуры, длившийся в течение юры и мела, т. е. около 130 млн лет. Факт остается фактом, видовые распределения меловых и юрских аммоноидей практически неразличимы. Интересно, что сильная положительная корреляция между сигмой и средним значением, выполнявшаяся в первые 200 млн лет эволюции аммоноидей, полностью исчезает в юрском и меловом периодах (см. рис. 23). Несостоятельность ортоселекции Выше в поиске рационального объяснения долговременных эволюционных тенденций мы остановились на двух гипотезах — ортоселекции и росте адаптируемости. Хотя ортоселекция на первый взгляд выглядит предпочтительнее (поскольку обладание более сложной структурой уже кажется благом), эта гипотеза не в состоянии объяснить феномен застоя. Если преимущества сложной лопастной линии ощущались в первые 200 млн лет эволюции аммоноидей, то они должны были сохранять свое значение и далее, но за последние 130 млн лет доля аммоноидей с довольно простой сутурой не убывала. Также трудно объяснить отсутствие прогресса в строении лопастной линии гониатитов середины карбона и цератитов конца триаса (см. рис. 22, б). Другой сильный аргумент против ортоселекции был представлен американским палеонтологом Дж. Цисне, проследившим за ростом дифференциации конечностей у водных членистоногих. Для количественной оценки этой формы морфофункционального прогресса Цисне предложил использовать степень тагмозиса конечностей (h), которую можно вычислить по формуле (2.1). Каждый отряд водных членистоногих (от примитивных трилобитов до высших ракообразных) был охарактеризован величиной h, после чего было проанализировано, как в течение 600 млн лет эволюции этих беспозвоночных изменялся средний для отряда уровень тагмозиса. Оказалось, что в ходе исторического развития водных членистоногих величина <h> возрастала, следуя S-образной (логистической) кривой (рис. 24). В течение первых 250 млн лет шло довольно быстрое увеличение среднего тагмозиса, затем этот процесс стал тормозиться и практически прекратился к началу мезозоя. Так что за последние 150 млн лет уровень <h> не повышался, хотя одни отряды вымирали, а другие возникали.
 Рис. 24. Рост среднего тагмозиса (<h>) конечностей водных членистоногих в историческом развитии (по: [Cisne, 1974]).
E — кембрий, О — ордовик, S — силур, Р — палеоген, N — неоген, R — современность. Остальные условные обозначения см. рис. 22.
Рис. 24. Рост среднего тагмозиса (<h>) конечностей водных членистоногих в историческом развитии (по: [Cisne, 1974]).
E — кембрий, О — ордовик, S — силур, Р — палеоген, N — неоген, R — современность. Остальные условные обозначения см. рис. 22.
Данное явление нельзя объяснить исчерпанием возможностей к прогрессивной дифференциации конечностей, так как средний тагмозис на стадии плато (<h> = 1,8) намного ниже, чем у десятиногих ракообразных (h = 2,5). Низкий уровень плато объясняется существованием в течение многих миллионов лет практически постоянной доли «примитивных» отрядов. Это плато продолжительностью в 150 млн лет явно свидетельствует против таинственных внутренних (гомогенетических) факторов, толкавших членистоногих по пути прогресса, но, самое главное, оно ставит серьезные трудности и перед сторонниками ортоселекции. Ведь выгода от функциональной дифференциации конечностей, казалось бы, очевидна. Тогда почему же доля примитивных отрядов не убывала в течение последних 150 млн лет? В 1949 г. Т. Уэстолл предпринял попытку измерить темп прогрессивной эволюции у двоякодышащих рыб. В данном случае уровень организации оценивался числом «прогрессивных» структурных новшеств (числом баллов). Динамика прогресса структурной сложности имеет тот же S-образный вид: период быстрого увеличения числа баллов продолжительностью около 150 млн лет сменяется гораздо более долгим плато (рис. 25).
 Рис. 25. Увеличение числа прогрессивных признаков (в баллах) в историческом развитии двоякодышащих рыб (по: [Рефф, Кофмен, 1986]).
Рис. 25. Увеличение числа прогрессивных признаков (в баллах) в историческом развитии двоякодышащих рыб (по: [Рефф, Кофмен, 1986]).
Характер кривых, передающих три случая морфологического прогресса (рост тагмозиса у членистоногих, сложности лопастной линии у аммоноидей и продвинутости организации у двоякодышащих рыб) весьма сходен. Во всех случаях первоначальный период подъема сменяется периодом застоя. Объяснение этому удивительному феномену мы попытаемся дать в следующей главе.
Глава 6. Мобилизующий отбор
Эволюция подотряда Ammonitina протекала необычайно бурно. Все морфологические особенности раковины — и внешняя форма, и скульптура, и перегородки — проявляли при этом сильнейшую мобильность. <…> Способы усложнения лопастной линии были также необычайно многообразны.Мутабильность К проблеме генетической сложности структуры можно подойти еще с одной стороны. Дело в том, что специальные гены, как и всякие другие, обязаны мутировать, порождая внутрипопуляционную изменчивость по мощности соответствующих рабочих структур. Заметим, что мощность морфологической структуры, с точки зрения генетика, следует отнести к мерным (количественным) признакам, для которых типичен непрерывный характер изменчивости. Мутации по этим признакам можно разделить на две четко очерченные категории — с качественным и количественным эффектом. Первые вызывают настолько резкое изменение значения признака, что наблюдатель видит их эффект без всяких измерений. Примерно также проявляет себя и потеря гена, поэтому в основе мутаций с качественным эффектом обычно лежит серьезное повреждение функции гена. Мутации второй категории ведут к небольшому (на несколько процентов) сдвигу величины признака, и этот эффект, как правило, меньше шума среды. Еще одна особенность данных мутаций заключается в том, что они могут не только снижать, но и повышать значение признака. Поэтому природные популяции нередко хорошо отвечают на отбор по мерным признакам в обоих направлениях. За мутациями этого типа и за возникающими в результате их действия аллелями исторически закрепилось несколько неудачное название полигенных. В гл.4 мы рассмотрели аддитивную модель наследования количественных признаков. Суть ее сводилась к тому, что величина признака складывается из суммы примерно равных вкладов большого числа генов. Обширный опыт биометристов показывает, что реальная картина наследования количественных признаков весьма часто начинает неплохо соответствовать аддитивной модели, если их измерять логарифмической, а не обычной (арифметической) шкалой. Особенно четко «улучшающий» эффект логарифмирования проступает, когда значение признака у родителей отличается в несколько раз. Столь большая разница обычно бывает у форм, разделенных или длительной эволюцией, или длительной селекцией. Следовательно, разность логарифмов величины признака родительских форм можно трактовать как сумму соизмеримых эффектов аллельных замещений по значительному числу локусов (см. (5.6)). Аддитивность логарифма отражает мультипликативность величины признака, измеренного арифметической шкалой. Иными словами, эта величина представима в виде произведения близких по значению сомножителей, за каждым из которых стоит отдельный ген. В предыдущем разделе мы показали, что мультипликативный характер генетического контроля размеров и мощности рабочей структуры возникает в результате весьма длительного движущего отбора, направленного на интенсификацию ее функции. При этом идет аккумуляция генов (названных нами специальными), управляющих программой развития структуры. Наши знания о скоростях полигенных мутаций базируются на данных, полученных при исследовании очень важного количественного признака — жизнеспособности. Мы уже знаем (гл.4), что мутации по этому признаку разделяются также на две категории: 1) рецессивные летали и полулетали, снижающие жизнеспособность не менее чем в 10 раз; 2) слегка вредящие мутации, снижающие в гомозиготном состоянии жизнеспособность всего на 2–4 %. Напрашивается явная аналогия с только что рассмотренным делением на две категории мутаций по обычным мерным признакам. Летали и полулетали подобны мутациям с качественным эффектом, а слегка вредящие мутации можно считать аналогами полигенных. Т. Мукаи так и назвал их «полигенными мутациями по жизнеспособности». Напомним, что этот японский генетик, следя за возникновением мутаций обеих категорий, сделал важное открытие — он установил, что у дрозофилы полигенные мутации по жизнеспособности возникают в 20–30 раз чаще, чем летали и полулетали. С эволюционной точки зрения наибольший интерес представляют именно полигенные мутации. Во-первых, они относительно безвредны, т. е. незначительно сказываются на основных показателях приспособленности, и, во-вторых, по скорости возникновения, по-видимому, не менее чем на порядок превосходят мутации с качественным эффектом. Молекулярная природа генов с количественным эффектом Здесь мы коснемся сложного вопроса, еще далекого от полной ясности, однако весьма немаловажного для последующего изложения, — молекулярной организации наследственного вещества. Хромосома многоклеточных — это очень длинная (до нескольких сантиметров) единая молекула ДНК. Хотя она и является носителем наследственной информации, из этого совсем не следует, что любая нуклеотидная последовательность генома обязательно влияет на приспособленность. Анализ строения ДНК многоклеточных показывает, что генетическая информация (по крайней мере, доступная нашему пониманию) размещена вдоль этой гигантской молекулы далеко не равномерно. Более того, нет сомнений, что гены занимают в ней только небольшую часть. При этом сам ген состоит из совершенно разных по информационному содержанию сегментов — экзонов и интронов. Первые чаще всего кодируют первичную структуру какого-нибудь белка, а вторые обычно ничего не кодируют (рис. 26). В межгенных промежутках (а иногда и в интронах) разбросаны нуклеотидные последовательности, имеющие отношение к генной регуляции. Активность гена проявляется в производстве им своего слепка — молекулы РНК, из которой затем вырезаются участки, соответствующие интронам. Заметим, что транскрипция РНК является лишь одним из этапов экспрессии гена. Другие этапы включают в себя синтез, сборку и созревание белковых молекул, а также их транспорт и встраивание в соответствующие субклеточные структуры. В многоклеточном организме большое число генов находится под жестким контролем программ развития всевозможных морфологических структур, и многие из них активны только в клетках определенного типа. Многочисленные данные, полученные в последнее время, свидетельствуют, что такой контроль осуществляется согласованным взаимодействием регуляторных белков (транс-факторов) с довольно короткими и весьма специфичными последовательностями ДНК (цис-элементами). Большинство цис-элементов расположено невдалеке от точки начала транскрипции гена; одни из них ответственны за тканеспецифичность генной экспрессии, другие — за ее уровень.Б.Е. Руженцев
 Рис. 26. Строение сегмента хромосомы эукариот. а — неактивный, б — активный ген. 1 — экзоны; 2 — интроны; 3 — межгенные участки; 4 — цис-элементы; 5 — транс-факторы; 6 — РНК-полимераза.
Рис. 26. Строение сегмента хромосомы эукариот. а — неактивный, б — активный ген. 1 — экзоны; 2 — интроны; 3 — межгенные участки; 4 — цис-элементы; 5 — транс-факторы; 6 — РНК-полимераза.
Для включения гена (т. е. для инициации транскрипции РНК) необходимо образование комплекса транс-факторов с белками транскрипционной машины (представленной прежде всего субъединицами РНК-полимеразы). При этом цис-элементы сближаются, а отрезки ДНК между ними сгибаются в петли. Таким образом, большой сегмент хромосомы с геном и протяженными (до нескольких тысяч нуклеотидов) соседними последовательностями выступает как единая матрица для сборки мультимолекулярного ансамбля, инициирующего транскрипцию. Понятно, что изменение числа цис-элементов и даже изменение расстояния между ними может отразиться на скорости сборки такого ансамбля, т. е. в конечном счете на уровне генной экспрессии. Хромосомы многоклеточных буквально начинены так называемыми мобильными элементами — всевозможными последовательностями ДНК, способными к независимому «размножению» и передвижению внутри генома. По-видимому, именно эти перемещения ответственны за возникновение большей части так называемых спонтанных мутаций, составляющих основу генетической изменчивости природных популяций. Легко представить, что встраивание довольно крупного отрезка чужеродной ДНК в экзонный участок гена приведет к инактивации последнего, а соответствующая мутация пополнит ряды рецессивных леталей, подлежащих удалению очищающим отбором. Однако гораздо чаще мобильные элементы должны попадать в протяженные межгенные промежутки, что может отразиться лишь на экспрессии расположенных по соседству генов. На уровне фенотипа такие изменения наследственного материала будут выглядеть скорее всего как полигенные мутации. Мукаи проследил за уровнем экспрессии гена алкогольдегидрогеназы (фермента, восстанавливающего спирт) у дрозофилы в течение 300 поколений. Хромосома с этим геном была выведена из-под контроля очищающего отбора, что позволяло проследить за накоплением спонтанных мутаций, затрагивающих активность фермента. За время наблюдения межлинейная дисперсия по ферментативной активности возросла в 27 раз, хотя среднелинейное значение активности осталось прежним. Также не изменились и физико-химические свойства алкогольдегидрогеназы, обусловленные первичной структурой белка. Мукаи предположил, что мутации затронули какие-то районы, расположенные за пределами гена. Мобильность По Дарвину, эволюция основана на отборе особей с едва заметными уклонениями фенотипа. Поскольку отбор направлен на изменение величины определенного количественного признака, принципиальное значение имеет не мутабильность генов, а мутабильность признаков. Существует несколько способов измерения последней. Например, каждая особь изогенной линии самоопыляющихся растений может стать основательницей дочерней сублинии. Если все особи-основательницы имеют тождественные генотипы, то средние значения количественных признаков в сублиниях будут совпадать. Если же особь-основательница несет мутацию, затрагивающую развитие одного из количественных признаков, то среднее значение данного признака у потомков мутантною индивида скорее всего будет смещено. Следовательно, определяя генотипические значения признака в большой совокупности дочерних сублиний, можно оценить уровень его мутабильности. Такого рода анализ был проведен у кукурузы и у мыши, в обоих случаях получена удивительно высокая величина мутабильности — около 0,01 мутаций на признак на геном за поколение. Таким образом, в результате мутирования в любой крупной популяции за одно поколение должна возникать новая генетическая дисперсия по любому количественному признаку. Получается, что даже изогенная линия может отвечать на отбор. Как мы помним (см. уравнение (4.19)), величина этого ответа пропорциональна наследуемости признака, т. е. доле генетической компоненты в его фенотипической дисперсии. В экспериментах по отбору на число щетинок в изогенных линиях дрозофилы скорость появления мутационной дисперсии оценена в несколько тысячных от величины фенотипической дисперсии. Длительный и интенсивный отбор должен вести к постепенному исчерпанию запасов генетической изменчивости в популяции. Поэтому ответ на отбор в конце концов будет определяться лишь скоростью возникновения мутационной дисперсии. Процесс мутирования по разным генам независим, поэтому в первом приближении можно считать, что мутационная дисперсия по величине признака, возникающая за поколение, равна сумме мутационных дисперсий по каждому локусу. Выходит, что с ростом генетической сложности признака должна расти и мутационная дисперсия его величины. В предыдущей главе мы установили, что увеличение числа специальных генов ведет к росту размеров и мощности рабочей структуры. В связи с этим приобретает особое значение вопрос, как будут соотноситься между собой обе характеристики структуры (мощность и мутационная изменчивость) при увеличении числа специальных генов. В качестве меры изменчивости структуры, обусловленной мутированием одного локуса, удобно взять сигму мутационного эффекта (σ0), которую можно трактовать как наиболее вероятное значение сдвига мощности структуры при мутировании одного из ее специальных генов. Так как естественный отбор «измеряет» эффект мутационных замен логарифмической линейкой, то имеет смысл оценивать величину σ0 в долях от мощности интактной структуры (напомним, что ΔlnP ≈ ΔР/Р). Если бы мощность органа определялась суммой вкладов его специальных генов, то величина относительного прироста мощности при добавлении к программе развития еще одного гена была бы тем меньше, чем выше генетическая сложность органа. Очевидно, что в данном (аддитивном) случае генетического контроля накопление специальных генов вело бы к созданию рабочих структур, мало чувствительных к мутациям, так как величина сигмы мутационного эффекта уменьшалась бы с увеличением числа их специальных генов. Однако обычно мощность органа представима в виде произведения сомножителей, олицетворяющих собой вклады отдельных специальных генов, поэтому добавление еще одного такого гена увеличивает мощность органа в некоторое число раз (вне зависимости от его мощности или от числа специальных генов), т. е. величина относительного прироста мощности рабочей структуры не зависит от ее генетической сложности. В данном (мультипликативном) случае генетического контроля увеличение числа специальных генов не снижает чувствительности органа к мутациям, поскольку любой сдвиг (в процентах) вклада отдельного гена представляет собой сдвиг на тот же процент мощности органа. Таким образом, сигма мутационного эффекта на мощность рабочей структуры не должна зависеть от числа ее специальных генов. Начиная с Дарвина неоднократно используется метафора, что естественный отбор как бы лепит новые формы, поэтому сам собою возник вопрос о пластичности материала, с которым имеет дело эволюция. Часто высказывалось, по-видимому, совершенно правильное мнение, что биологический материал не пластилин, что формирование структур подчиняется своим законам и т. д. Не входя в дискуссию о «законах» образования биологических структур, заметим, что с функциональной точки зрения имеет значение только их мощность. В связи с этим вместо несколько неопределенной пластичности биологического материала лучше говорить об эволюционной изменчивости мощности рабочих структур. Эту изменчивость назовем мобильностью и будем ее оценивать по скорости, с которой под действием длительного движущего отбора изменяется мощность структуры. А учитывая «логарифмические очки» естественного отбора (т. е. его «склонность» оценивать не абсолютный, а относительный сдвиг величины признака), назовем мобильностью скорость, с которой изменяется логарифм мощности рабочей структуры под действием длительного движущего отбора определенного давления. Еще раз напомним, что небольшой относительный сдвиг величины признака (измеренный в долях от его исходного значения) можно трактовать как сдвиг логарифма величины признака. Как ответ на длительный отбор мобильность (М) должна прямо зависеть от скорости возникновения мутационной дисперсии логарифма мощности структуры, что, в свою очередь, определяется числом специальных генов (N), их средней мутабильностью и средней дисперсией мутационного эффекта <σ02>. Поскольку мощность структуры определяется мультипликативным действием ее специальных генов, то сигма мутационного эффекта σ0, измеренная в долях от мощности интактной структуры, не зависит от числа этих генов. Кроме того, число специальных генов любой морфологической структуры довольно велико, поэтому значение <σ02> для разных структур, по-видимому, различается незначительно. Наконец, по средней мутабильности специальных генов рабочие структуры также не могут существенно различаться. Таким образом, с достаточной точностью должно выполняться соотношение М ~ N, (6.1) т. е. мобильность структуры пропорциональна числу ее специальных генов. Перевернув назад несколько страниц, можно убедиться в том, что совсем недавно мы установили почти тривиальную связь между мощностью структуры у среднестатистического вида филетической группы, с одной стороны, и числом его специальных генов — с другой (см. (5.10)). Сопоставив это соотношение с (6.1), получаем, что <lnP> ~ <М>, (6.2) т. е. в пределах филетической группы средневидовой логарифм мощности рабочей структуры пропорционален ее средневидовой мобильности. Дифференциальное выживание филетических линий Представим себе совокупность родственных видов, обитающих в сходных экологических условиях. Пусть все особи этих видов снабжены набором гомологичных рабочих структур. Отличия касаются лишь их размеров и деталей строения. Вообразим, что в среде обитания данной таксономической группы начались однонаправленные изменения какого-то жизненно важного фактора. В таких условиях мощность одной из рабочих структур может стать недостаточной для сохранения прежней численности популяций. Теперь любой вид должен успеть скомпенсировать ухудшение среды усилением мощности лимитирующей структуры, иными словами, судьба вида попадает в зависимость от скорости усиления этой структуры. Если данная скорость не сможет затормозить процесс падения абсолютной приспособленности вида, то он вымрет. Пусть перед каждым видом поставлена одна и та же задача — увеличить мощность гомологичной структуры на один и тот же процент (скажем, на 30 %). Быстрее всего эту задачу можно решить, усилив экспрессию нескольких специальных генов, но для этого вид должен успеть за отведенный ему судьбою срок зафиксировать несколько благоприятных мутаций. Темп фиксации последних при прочих равных условиях пропорционален скорости их возникновения, а она тем выше, чем больше генов участвует в контроле развития структуры. Таким образом, мы получаем удивительный результат: чем сложнее устроена лимитирующая структура, тем больше шансов у обладающего ею вида на филетическое выживание, и, наоборот, чем проще она устроена, тем больше у вида шансов на вымирание. Напомним, что для равенства всех прочих условий степень благоприятности мутаций не должна зависеть от числа специальных генов, а это выполняется при мультипликативном характере генетического контроля. Генетически сложная структура может быстро изменить свою мощность и в противоположном направлении, т. е. снизить ее. Для этого достаточно ослабить экспрессию нескольких специальных генов. Поэтому сложно устроенная структура сообщает виду повышенные шансы на выживание даже в ситуации, когда среда как бы благоприятствует более примитивным. Вид — обладатель генетически примитивной структуры не может быстро сделать ее еще примитивнее, так как слишком мало генов контролирует ее развитие. Таким образом, вне зависимости от того, чего требует окружающая среда от рабочей структуры — усиления или ослабления — главное, что определяет судьбу вида, заключается в скорости изменения мощности лимитирующей структуры. Во всех случаях больше шансов на выживание имеют виды, у которых такая структура управляется большим числом генов. Это обстоятельство можно выразить в терминах более обычных для теории эволюции: на виды, эволюционирующие в одной адаптивной зоне, постоянно действует отбор, благоприятствующий увеличению генетической сложности структур, от которых среда регулярно требует усиления или ослабления функции. Такой отбор имеет смысл назвать мобилизующим, так как главная «цель», которую он преследует, заключается в увеличении мобильности структур, наиболее часто лимитирующих скорость адаптации. Итак, мобилизующий отбор ведет к постоянному увеличению числа специальных генов лимитирующей структуры. Откуда эти гены берутся? Создать совершенно новый ген за короткое время невозможно. Может быть где-то в геноме хранится запас готовых к употреблению генов? Однако такого запаса нет и быть не может, так как если ген не работает, то он не проверяется на качество очищающим отбором. Аллельный состав такого гена подчиняется только законам нейтральной эволюции, поэтому под действием мутационного давления через несколько миллионов лет все локусы генного резерва были бы замещены бессмысленными аллелями, превратились бы, как говорят, в хлам из ДНК. Однако для увеличения числа специальных генов, контролирующих развитие лимитирующей структуры, можно воспользоваться вполне исправными генами, которые в данный момент обслуживают другие структуры. Такой ген (из сферы обслуживания других функций) можно было бы удвоить (дуплицировать), а его дополнительную копию, снабдив новыми контролирующими элементами, направить в распоряжение лимитирующей структуры. Хотя этот путь вполне реален, у него есть один недостаток — он ведет к увеличению наследственной информации генома, а следовательно, и к росту мутационного генного груза. Конечно, добавление одного лишнего гена — пустяк, и очищающий отбор этого даже не заметит. Но надо полагать, объем наследственной информации генома был доведен до предельной черты еще на заре эволюции многоклеточных. Мы уже упоминали о гипотезе А.С. Кондрашова, по которой даже разделение особей на два пола, несмотря на все связанные с этим энергетические и информационные издержки, служит главным образом цели снижения мутационного груза. Таким образом, генные дупликации как стратегическое направление едва ли перспективны. Куда изящнее выглядит просто смена геном своей специализации (его перепрофилирование), когда ген, управлявший ранее развитием одной структуры, после соответствующей мутации начнет повышать мощность другой, лимитирующей, структуры. Кроме того, вполне вероятно, что в измененной среде мощность некоторых структур может оказаться избыточной. На первый взгляд, это может показаться странным, но достаточно вспомнить о видах, заброшенных эволюционной судьбой в подземные пещеры. Многочисленные структуры зрительного анализатора, покровительственная окраска, формы поведения, рассчитанные на зрительное восприятие, — все это в полной темноте становится избыточным, излишним. Временная избыточность структур открывает путь к перепрофилированию их специальных генов. Развивая эту мысль, можно допустить, что изменение среды ведет к «перекачке» генов из системы обслуживания структур с избыточной мощностью в состав специальных генов лимитирующей структуры. Генная специализация Организмпредставляется нам совокупностью взаимодействующих друг с другом морфологических рабочих структур, при этом на клетки мы смотрим как на элементарные единицы нескольких типов, которые могут размножаться и производить молекулярные продукты, нужные не им самим, а организму в целом. Такими продуктами могут быть и пищеварительные ферменты, и гормоны, и эмбриональные индукторы. Поскольку функция генов, управляющих синтезом этих продуктов, заключается в создании над клеточного уровня организации, мы можем назвать их генами межклеточного взаимодействия. В то же время нам хорошо известно, что клетки далеко не атомы. Для реализации основных клеточных процессов — репликации ДНК, синтеза и созревания РНК, синтеза белков, производства и запасания энергии, работы митотического аппарата и многого другого — задействованы тысячи генов. Их принято называть генами домашнего хозяйства. Таким образом, все гены многоклеточного организма можно разделить на два обширных класса — гены межклеточного взаимодействия, ответственные за создание надклеточных структур, и гены домашнего хозяйства, нужные для работы любой клетки. Гены с новой молекулярной функцией появляются в процессе эволюции очень редко, поэтому рядовой эволюционный сдвиг межвидового ранга связан не с появлением качественно новых генов, а с изменением времени, места и, главное, уровня экспрессии уже имеющихся. Для увеличения числа специальных генов необходимо выполнение двух условий. Во-первых, какой-нибудь «посторонний» ген нужно снабдить новым цис-элементом, чтобы он мог работать в клетках, формирующих рабочую структуру, во-вторых, перепрофилированный ген должен включиться в программу развития лимитирующей структуры, т. е. он увеличит ее мощность. Правда, события подобного рода не могут происходить часто, так как в окрестности специального гена должна появиться вполне конкретная, хотя и короткая, последовательность ДНК. Вместе с тем уже простого изменения взаимного расположения цис-элементов вблизи кодирующих последовательностей может хватить для ощутимого сдвига уровня генной экспрессии. Очевидно, что такого рода мутации могут происходить не так уж редко. Однако какой бы малой ни казалась нам вероятность перепрофилирования локусов, прямой анализ строения генов, обслуживающих морфологические структуры, часто указывает на их происхождение от генов домашнего хозяйства. Особенно впечатляют в этом отношении белки хрусталика глаза — кристаллины. α-кристаллины явно родственны белкам теплового шока, а ε-кристаллин птиц и рептилий фактически является одной из форм фермента метаболизма глюкозы — лактатдегидрогеназы. Основные функции многоклеточного организма в значительной мере обеспечиваются его надмолекулярными структурами. Каждая из них возникает в ходе индивидуального развития под контролем своих специальных генов, работа которых так тонко увязана между собой (в программу развития), что изменение экспрессии любого из них немедленно отражается на функции взрослой структуры. Фактически это означает, что возникающие с повышенной частотой мутации по генной экспрессии начинают выступать в роли мутаций, ослабляющих или усиливающих мощность рабочих структур. Отметим, что изменения наследственного материала, ведущие к увеличению числа специальных генов надклеточных рабочих структур, можно с полным основанием отнести к мутациям, повышающим «выход» благоприятных мутаций. Математическая модель мобилизующего отбора Попробуем более формально проанализировать механизм действия мобилизующего отбора. Представим себе, что в какой-то адаптивной зоне обитает большое число родственных видов, обладающих одним и тем же признаком — гомологичной и изофункциональной рабочей структурой. Пусть величина этого признака (под которым в данном случае мы понимаем логарифм мощности структуры) определяется суммарным вкладом его специальных генов. Каждый из них может находиться в двух аллельных состояниях 1 и 0. Аллель 1 соответствует активному состоянию гена (когда он экспрессируется), аллель 0 — отсутствию экспрессии. Таким образом, величина признака (Z) определяется числом специальных генов, находящихся в активном состоянии. Полное число генов (G) в геноме разных видов постоянно, тогда как число специальных генов (N), определяющих степень развития признака, различно. Гены, не вошедшие в состав специальных (их число равно G — N), будем называть «сторонними». Они определяют развитие каких-то других (сторонних) признаков. В среде обитания время от времени происходят медленные однонаправленные изменения, когда над каждым видом нависает реальная угроза гибели. Избежать вымирания вид может за счет компенсаторного изменения величины признака на какое-то число единиц. (Что соответствует увеличению мощности рабочей структуры в какое-то число раз.) Увеличение признака на одну единицу может произойти за счет фиксации мутаций двух типов. Во-первых, это могут быть мутации, активирующие специальные гены, и, во-вторых, мутации, переводящие ген из разряда «сторонних» в разряд специальных (с аллельным состоянием 1). Точно так же сдвиг признака в сторону его уменьшения может быть достигнут либо инактивацией одного специального гена, либо переводом активного специального гена в состав «сторонних». Для своего филетического выживания каждый вид должен за единицу макроэволюционного времени осуществить фиксацию нескольких благоприятных мутаций (любого типа), изменяющих признак на требуемую величину ΔZ. Донорами этих мутаций являются чаще всего специальные гены признака, так как изменение экспрессии гена — более вероятное событие, чем его перепрофилирование. Соответственно вероятность филетического выживания вида (его адаптируемость) будет расти с увеличением числа специальных генов (N). Это создает предпосылки роста генетической сложности признака. Такая простая модель позволяет осуществить машинный эксперимент. На каждом шаге модели в окружающей среде каждого вида происходят случайные изменения, которые никак не связаны с изменением среды обитания остальных видов. От одних видов среда требует увеличения признака, а от других — уменьшения. Всю эту картину можно передать типом распределения величины ΔZ. В данном эксперименте мы остановились на биномиальном симметричном распределении. С подробностями вычисления вероятностей выживания видов в новых условиях среды можно ознакомиться в книге «Основные факторы макроэволюции».
 Рис. 27. Моделирование динамики видовых распределений по величине признака (а) и числу его специальных генов (б). Приведенные распределения (слева направо) соответствуют 20, 50, 100, 200, 800-му шагу модели. Последнее распределение финальное.
Рис. 27. Моделирование динамики видовых распределений по величине признака (а) и числу его специальных генов (б). Приведенные распределения (слева направо) соответствуют 20, 50, 100, 200, 800-му шагу модели. Последнее распределение финальное.
Проследим за изменением видовых распределений по величине признака (рис. 27, а). В начальный момент все виды обладают минимальным значением признака, поскольку его развитие обеспечивает всего один специальный ген. Однако завершается машинная эволюция широким финальным распределением со средним значением, равным 18,4. Таким образом, несмотря на то, что среда с равной вероятностью благоприятствует как увеличению, так и уменьшению значения признака, его средневидовая величина (<Z>) неуклонно растет. Правда, параллельно с этим нарастает и уровень межвидовых различий по степени развития признака (σz) (рис. 28). В итоге даже в финальном распределении после долгой эволюции сохраняется довольно много видов с «архаичной», «примитивной», величиной признака. По-иному ведет себя распределение видов по числу специальных генов (рис. 27, б). По скорости роста среднего значения оно значительно опережает рассмотренное выше распределение по величине признака. Напомним, что рост числа специальных генов отражает усложнение генетической программы развития признака. Своеобразие динамики распределения видов по числу специальных генов состоит в том, что его размах стабилизируется гораздо раньше, чем среднее значение; поэтому в течение длительного отрезка времени мы наблюдаем практически параллельный снос вправо всего распределения. Следовательно, аккумуляция специальных генов идет со сходной скоростью во всех филетических линиях нашей эволюционирующей группы. Финальное распределение соответствует крайней специализации видов в отношении данного признака, поскольку большая часть генов генома попадает в разряд специальных (их среднее число равно 36,2). Ясно, что завершение процесса специализации обусловлено исчерпанием возможностей генома, число генов которого в данной модели неизменно (G=50).
 Рис. 28. Изменение среднего значения (<Z>) и сигмы (σ) распределений, представленных на рис. 27.
Рис. 28. Изменение среднего значения (<Z>) и сигмы (σ) распределений, представленных на рис. 27.
Так как мобильность признака определяется прежде всего числом специальных генов, то рост их среднего числа можно интерпретировать как рост мобильности признака. Мы видим, что модель вполне адекватно передает суть мобилизующего отбора. Сравнивая оба типа финальных распределений (по N и по Z), следует отметить, что виды «архаичные» и «примитивные» в отношении величины признака оказываются вполне «продвинутыми» по его генетической сложности. Кажущаяся архаичность, очевидно, обусловлена низким уровнем экспрессии большинства специальных генов. Однако мобильность признака у этих «псевдоархаичных» видов довольно высока, поэтому, если начать эволюцию именно с них, финальное распределение достигается гораздо быстрее. При сравнении данных модельных распределений с реальными распределениями аммоноидей по сложности сутуры нельзя не отметить их разительного сходства, которое распространяется даже на пропорциональность среднего значения и сигмы. Все это позволяет нам предположить, что сущность прогресса лопастной линии состояла в увеличении ее мобильности и, по-видимому, эта цель была достигнута аммоноидеями к началу юры, когда они «освоили» все способы рассечения сутуры. Принцип максимальной адаптируемости За долгие 700–800 миллионов лет истории многоклеточных предки каждого современного вида прошли через сотни (а может быть, и тысячи) актов проверки на адаптируемость. В связи с этим на каждую особенность строения современных организмов мы можем взглянуть как на приспособление, повышающее шансы вида на филетическое выживание. Иными словами, организация современных живых существ в какой-то степени должна отвечать принципу максимальной адаптируемости, т. е. максимальной скорости фиксации благоприятных мутаций. Рассмотрим филетическую группу гипотетических организмов, обладающих всего двумя функционально независимыми рабочими структурами А и Б. Во внешней среде временами случаются два типа изменений: один «требует» усиления (или ослабления) мощности структуры А, а другой предъявляет аналогичные требования к структуре Б. Хотя эти изменения происходят совершенно случайно, их можно охарактеризовать своими частотностями (вероятностями). Например, изменения первого типа могут случаться в 2 раза чаще, чем второго, тоща суммарное время, в течение которого мобилизующий отбор «увеличивает» число специальных генов структуры А, будет в 2 раза больше, чем время, когда он «добавляет» гены к структуре Б. Если число генов в геноме постоянно, то через некоторое время между специальными генами обеих структур должно установиться динамическое равновесие в соответствии с вероятностями требований к ним среды. При этом структура А «перетянет» на себя в 2 раза больше генов, чем структура Б. Заметим, что именно специальные гены и являются основными поставщиками благоприятных аллелей для движущего отбора, когда среда «требует» изменения той или иной структуры. Очевидно, что по «выходу» благоприятных мутаций структура A в 2 раза превосходит структуру Б, точно отражая соотношение численностей их специальных генов (а стало быть, и мобильностей). Иными словами, при одном и том же давлении отбора на обе структуры благоприятные мутации по структуре А будут возникать и фиксироваться вдвое чаще, чем по структуре Б. Теперь снабдим наши гипотетические организмы большим числом функционально независимых структур, при этом будем считать, что адаптивная зона предъявляет к каждой из них требования со своей характерной частотой. Совершенно ясно, что после длительной эволюции между численностями специальных генов отдельных структур установятся равновесные соотношения, которые будут соответствовать вероятностям требований среды к изменению их мощности. Фактически это можно трактовать как приобретение организмами знаний о своей адаптивной зоне. Филетическая линия как бы изучает среду обитания и приобретенные знания воплощает в генетическую сложность морфологических структур. Чем чаще структура лимитировала адаптацию в прошлом, тем она сложнее, тем выше выход благоприятных мутаций при отборе, направленном на ее изменение, и тем легче будет вид преодолевать в грядущем изменения среды, типичные для его адаптивной зоны. В итоге максимальной мобильности достигает орган, чья мощность чаще всего оказывается недостаточной (или избыточной) для проникновения вида в соседнюю экологическую нишу. Это означает, что в филетических линиях, вполне освоивших свое экологическое пространство, выход благоприятных мутаций достигает максимума. Более того, сам характер генного контроля мощности рабочих структур (мультипликативный) отвечает принципу максимальной адаптируемости. Во-первых, эффект мутации по любому специальному гену одинаково хорошо заметен для отбора и, во-вторых, величина этого эффекта не зависит от числа специальных генов. Такой тип генного контроля, очевидно, является следствием длительного движущего отбора, способного «видеть» лишь, во сколько раз (т. е. на какой процент) рабочая структура у особи с благоприятным аллелем развита сильнее (или слабее), чем у остальных членов популяции (см. гл.5). Повышенная чувствительность структуры к мутациям достигается при подчинении ее специальных генов единой программе развития, когда активность более поздних генов определяется активностью более ранних. Таким образом, привычный для нас характер возникновения структур в ходе онтогенеза соответствует принципу максимальной адаптируемости. Мобилизующий отбор в простой среде Мы всегда склонны считать окружающий мир сложным. Выражение «жизнь сложна» воспринимается нами как аксиома. Однако посмотрим на нашу жизнь попристальнее. За выполнение довольно простой и монотонной работы мы получаем деньги, которые затем обмениваем на пищу и кров. Кроме того, существующая плотность населения вполне достаточна для гарантии случайной встречи с особью другого пола. Теперь посмотрим, как решал те же проблемы первобытный человек. Попробуйте-ка выследить оленя и застрелить его из лука или найти в лесу съедобные растения и прокормить ими себя и своих детей. Да и спутника жизни обычно приходилось искать за тридевять земель на территории, занятой другим, нередко агрессивно настроенным родом. Сколько опасностей нужно было избежать при решении всех этих задач. Ясно, что жизнь современного человека намного проще. Адаптивные зоны филетических групп также различаются по степени сложности. В простых зонах повышенные требования предъявляются к одной из основных функций, поэтому действие мобилизующего отбора направлено на специализацию генов в отношении ограниченного круга рабочих структур. В итоге данные структуры усложняются, а большинство других упрощается (деградирует). Такая эволюционная стратегия традиционно называется специализацией и не считается прогрессом, хотя, с чисто структурной точки зрения, некоторые органы достигают удивительной сложности. Повышенная мобильность небольшого числа структур обеспечивает соответствующие филетические группы высоким темпом ветвления в простых адаптивных зонах. Беда только, что сами эти зоны не отличаются долговечностью, а с их исчезновением гибнет и все многообразие специализированных форм. По-видимому, именно эта стратегическая обреченность специализации не позволяет нам признать ее структурные успехи за прогресс. В отличие от прогресса проблема деградации привлекает к себе меньше внимания, что, вероятно, связано с оптимистичностью нашей натуры. Под деградацией обычно понимают вторичное упрощение рабочих структур. Так, переход к паразитизму ведет к ликвидации большинства структур, ранее обслуживавших функции доставки, удаления, гомеостаза и охраны. Червь, проникший в наш кишечник, получает доступ к уже переваренной пище. Ему не надо бояться хищников, поскольку это наша забота. От него же требуется только защититься от пищеварительных ферментов с помощью специальной кутикулы, прикрепиться с помощью крючков и присосок к стенке нашего кишечника и все. Единственной неудовлетворенной потребностью остается размножение. Правда, структуры, призванные для разрешения этой проблемы, достигают здесь необыкновенной сложности. Важнейшим приспособлением к паразитическому образу жизни обычно является колоссальная плодовитость. Так, у трематод мало того, что взрослые черви производят огромное число яиц, но и их личинки, проходя несколько стадий, могут на любой из них размножаться бесполым путем. При этом нередко каждая личиночная стадия протекает только в организме специфичного промежуточного хозяина. В то же время, если среда обитания вида-хозяина стала изменяться катастрофическим образом, то виды-паразиты должны успеть покинуть его (как крысы тонущий корабль) и подобрать себе другого хозяина. Адаптируемость для них — это прежде всего легкость смены видов-хозяев. Поэтому «деградацию» в данном случае можно рассматривать как колоссальный прогресс функции воспроизведения, «оттянувшей» на себя большую часть генов межклеточного взаимодействия. Эффективность такой стратегии демонстрирует и таксономический успех паразитов. Например, тип плоских червей насчитывает около 12,5 тысячи видов, из которых свободный образ жизни в сложных средах ведет всего около трех тысяч, остальные (9,5 тысячи) — паразиты. Мобилизущий отбор в сложной среде Иная судьба складывается у филетических групп, эволюционирующих в сложных адаптивных зонах, где регулярно предъявляются требования к гораздо большему числу рабочих структур. В этих условиях мобилизующий отбор не может обеспечить одну из них большим числом специальных генов, поэтому рабочие структуры чаще всего сохраняют «примитивный», «архаичный» вид. Эволюцию таких форм нельзя назвать бурной, их скромные структурные успехи теряются на фоне ярких достижений специализированных филетических групп. Зато овладение сложной средой обеспечивает архаичные (неспециализированные) формы весьма продолжительным существованием. Вместе с тем для этих «прозябающих» неспециализированных групп существует одна возможность — под давлением мобилизующего отбора резко поднять уровень адаптируемости. Она состоит в объединении специальных генов двух (или большего числа) рабочих структур для контроля развития одной новой, мобильность которой после такого объединения должна существенно возрасти. Данные сравнительной морфологии свидетельствуют о весьма широком распространении эволюционной тенденции к созданию морфологически сложных (составных) органов, возникших после интеграции нескольких более простых.
 Рис 29. Зубы акулы (по: [Оммани, 1975]).
Чешуи (а), покрывающие тело, заходя на поверхность челюстей, трансформируются в зубы (б).
Рис 29. Зубы акулы (по: [Оммани, 1975]).
Чешуи (а), покрывающие тело, заходя на поверхность челюстей, трансформируются в зубы (б).
Взять, к примеру, такое важнейшее для эволюции позвоночных событие, как появление челюстей — принципиально нового пищедобывающего органа. Это приобретение позволило рыбам «оторваться» ото дна и приступить к завоеванию всей водной толщи, поскольку челюсти открыли им доступ к новым источникам пищи. У первичных позвоночных глотка была прорезана серией парных щелей, на краях которых располагались жабры. Каркасом для этого дыхательного органа служила система висцеральных дуг — опорных элементов, размещенных в промежутках между жаберными щелями. Челюсти возникли при объединении скелета двух передних висцеральных дуг с прочным и относительно неподвижным мозговым черепом. Элементы одной висцеральной дуги преобразовались в верхнюю и нижнюю челюсти, тогда как другая висцеральная дуга взяла на себя роль «подвеска». Третьим структурным компонентом нового органа стали зубы, вначале представлявшие собой не что иное, как слегка измененную чешую, перешедшую на ротовую поверхность челюстей (рис. 29). Итак, мы видим, что данный орган удержания и умерщвления жертвы возник на основе по меньшей мере трех совершенно разных в функциональном отношении структур. Более всего нашему интуитивному представлению о морфологическом прогрессе отвечают составные полифункциональные органы, определяющие интенсивность сразу нескольких основных функций. Например, ясно, что органы зрения помогают и добыванию пищи, и защите от хищников, и поиску партнера по спариванию. То же верно для многих других структур, включая и такие сложные, как локомоторный аппарат, ответственный за движение животного. По большому счету, в данный аппарат помимо костей, мышц, суставов и сухожилий мы должны включить соматическую нервную систему и все органы чувств. Фактически все это составляет большую часть тела животного. Объяснение правила Копа Внешняя среда, несмотря на всю свою сложность, «интересует» вид в отношении очень небольшого числа вопросов: 1) есть ли в среде источники пищи для поддержания достаточно плотных популяций; 2) не слишком ли высок уровень истребления со стороны хищников и прочих биотических факторов; 3) не препятствуют ли физико-химические условия среды успешному завершению индивидуального развития. Если вид способен существовать в течение многих сотен поколений, значит, он получает удовлетворительные ответы на все эти вопросы. Также ясно, что в условиях медленно ухудшающейся среды один из перечисленных вопросов может остаться без ответа. Какова будет адаптационная реакция вида? Много ли у него путей выхода из кризиса? Вообще для восстановления уровня абсолютной приспособленности существует несколько путей. Эволюционирующий вид их не выбирает, а идет сразу по многим, но к цели (т. е. к спасению) ведет кратчайший. Длина же пути определяется скоростью появления полезных мутаций и степенью их благоприятности. Ситуация 1a. Поток пищи (энергии) ослабел по вине вида-конкурента. Существуют три варианта выхода. А. Усилить пищевой поток. Это возможно, если вид обладает развитой и мобильной пищедобывающей структурой. Данный вариант можно было бы назвать специальным. Б. Сохранить прежнюю биомассу при меньшей численности, т. е. увеличить размеры особи. В. Сохранить прежнюю численность при меньшей биомассе. Для этого достаточно уменьшить массу средней особи, что сопряжено с усилением удельного метаболизма и укорочением времени генерации. В итоге вид будет быстрее поглощать пищевой ресурс, чем может поставить конкурента на грань катастрофы. Ситуация 1б. Поток пищи ослабел в связи с уменьшением ресурса ниши. Параллельно в соседних нишах произошло примерно то же самое. Решение в данном случае может быть связано только с измельчанием особей при усилении функции воспроизведения. Ситуация 2. Повысился уровень гибели особей от хищников. Здесь можно указать на четыре варианта выхода из положения. А. Уйти от конкретного хищника, выработав специфические средства защиты (яд, шипы, панцирь и т. д.), — это специальный ответ. Б. Увеличить размеры особи, став хищнику, что называется, не по зубам. В. Скомпенсировать потери от хищника, снизив энергетические расходы единицы биомассы. Этого можно достичь, увеличив массу особи. Г. Скомпенсировать потери от хищника ускорением развития и усилением функции воспроизведения, что достигается уменьшением массы особи. Ситуация 3. Ухудшились физико-химические условия среды, что привело к замедлению роста и развития особей. Здесь снова может быть два типа адаптивного ответа — специальный и общий. При специальном ответе защита от неблагоприятного фактора заключается в изменении мощности конкретной рабочей структуры. Например, снижение температуры среды можно скомпенсировать или увеличением теплопроизводства (дрожание мышц у насекомых), или уменьшением теплоотдачи (подкожный жир, мех и пух у теплокровных животных). Однако ту же проблему можно решить, увеличивая (в соответствии с правилом Бергмана) размеры особей, поскольку это приведет к возрастанию отношения массы тела к его поверхности. Итак, мы приходим к заключению, что практически любой долговременный неблагоприятный для зоологического вида сдвиг среды может быть скомпенсирован или адекватным изменением мощности какой-то специализированной рабочей структуры, или изменением размера тела. Следовательно, размер особей можно рассматривать как своеобразный регулятор, способный изменять интенсивность фактически всех основных функций. В таком случае изменчивость массы индивида должна стать одной из главных «забот» мобилизующего отбора. Заметим, что увеличение массы животного предполагает аллометрический рост его одно- и двухмерных рабочих структур. Поэтому мобильность размеров тела попадает в зависимость от мобильности органов. Таким образом, мобилизующий отбор, «стремясь» к повышению пластичности размеров особи, обусловливает макроэволюционную тенденцию к увеличению числа специальных генов многих соматических структур, составляющих большую часть тела индивида. Это может объяснить тенденцию, описываемую правилом Копа. Откуда берутся гены для контроля соматических структур, если общее число генов в геноме неизменно? Мы уже упоминали, что у животных увеличение размеров тела уменьшает энергетическую стоимость единицы биомассы и, в частности, усредненной клетки организма. Это обстоятельство означает снижение требований к регуляции генов домашнего хозяйства, т. е. определенное снижение объема наследственной информации, обслуживающей внутриклеточные функции. Данный информационный выигрыш может быть использован для увеличения числа генов межклеточного взаимодействия. Кроме того, часть генов для контроля соматических органов можно получить «перекачкой» из сферы обслуживания функции воспроизведения, что должно сопровождаться прогрессивным уменьшением относительных размеров гонад и других репродуктивных структур. Мобильность лопастной линии аммоноидей Процесс возникновения новых форм в ходе эволюции весьма необычен, поэтому ему трудно подобрать какую-то аналогию из нашей практической жизни. Обратимся к миру фантазии. Вообразим, что мы летим в звездолете, и расстояние, которое предстоит преодолеть, столь велико, что на это путешествие должна уйти почти вся жизнь. Самое неприятное в этих условиях — непредвиденные задержки, но, к сожалению, именно с ними нам и пришлось столкнуться. Вопреки ожиданиям, межзвездное пространство оказалось неоднородным, и мы то и дело попадаем в зоны с аномальными, заранее непредсказуемыми свойствами. Выяснилось, что пройти эти зоны можно лишь при условии изменения кораблем своей формы, т. е. перед входом в каждую новую аномальную область пространства мы должны перестраивать оболочку звездолета, придавая ей вид (в зависимости от «требований среды») то диска, то спирали, то шара и т. д. На подобные операции требуется какое-то время, поэтому успех всего путешествия попадает в зависимость от скорости, с которой мы способны изменить форму корабля. Эту скорость мы могли бы назвать мобильностью оболочки; очевидно, что именно она и является главным (ведущим) свойством, от которого зависят наши шансы дожить до конца пути. Оболочка корабля, конечно же, состоит из разнородных частей (компонентов), которые различаются по назначению, конструкции, материалу, а стало быть, и по мобильности. Если функция данных частей допускает несколько конструктивных решений, то для ускорения полета естественно выбрать те, которые позволят перестроить корабль за минимальное время. При этом свои основные усилия мы сосредоточим на мобилизации наименее пластичных компонентов. Ясно, что после серии таких реконструкций мы снабдим звездолет оболочкой, все компоненты которой будут отвечать принципу максимальной мобильности. Теперь перейдем к эволюции аммоноидей. Сначала отметим, что ведущим признаком, изменение которого позволяет виду покинуть старую экологическую нишу или захватить новую, является размер особей, который для аммоноидей удобно оценивать по величине радиуса раковины. Мобильность сутуры, по существу, является только одной из компонент мобильности размера раковины. Другими ее компонентами могут выступать мобильности таких признаков раковины, как толщина стенки, число поперечных перегородок, наружная скульптура и др. Сдвиг величины логарифма радиуса раковины (ΔlnR), сопровождающий видообразование, очевидно, возникает как результат серии фиксаций благоприятных мутаций. Скорость фиксации каждой такой мутации определяется при прочих равных условиях селективным выигрышем, который получает ее носитель (см. (3.32)). Пусть мы имеем два класса мутаций, любая из которых укрепляет стенку раковины в одинаковой степени, но мутации первого класса утяжеляют раковину, а мутации второго класса — нет. Очевидно, что движущий отбор отдаст предпочтение мутациям второго класса из-за более высокого коэффициента селекции. Наиболее эффективный способ упрочить раковину, не изменяя массы (см. гл. 5), состоит в удлинении лопастной линии за счет увеличения числа ее изгибов, т. е. за счет повышения сложности. Следовательно, виды аммоноидей с более сложной сутурой имеют больше шансов на филетическое выживание, когда среда «требует» изменения размеров раковины. Данное обстоятельство способно обусловить макроэволюционную тенденцию к повышению мобильности лопастной линии. Мы регистрируем эту тенденцию как постепенное увеличение в ходе исторического развития среднего значения и размаха видовых распределений по сложности сутуры аммоноидей. В конце концов мобилизующий отбор мог бы привести к снабжению каждого вида сверхгибкой лопастной линией, способной адекватно изменять степень своей изогнутости при любом требовании среды к изменению величины и прочности раковины.
 Рис. 30. Динамика изменения среднеквадратического отклонения (сигмы) видовых распределений аммоноидей по размеру раковины σlnR и сложности лопастной линии σz.
Рис. 30. Динамика изменения среднеквадратического отклонения (сигмы) видовых распределений аммоноидей по размеру раковины σlnR и сложности лопастной линии σz.
Теперь попробуем оценить величину межвидовой изменчивости по сложности сутуры в филетической группе, где все виды обладают сверхгибкой лопастной линией. Площадь боковой поверхности раковины растет пропорционально квадрату ее радиуса и по тому же закону должна возрастать длина сверхгибкой сутуры (l∞). В то же время длина контура поперечного среза раковинной трубки (l0) пропорциональна ее радиусу (R), поэтому отношение l∞/l0 тоже должно быть пропорционально радиусу, т. е. l∞/l0 = kR, (6.3) где k — коэффициент пропорциональности. В логарифмической форме равенство (6.3) преобразуется в ln(l∞/l0) = lnR + lnk. (6.4) Заметим, что в левой части этого уравнения стоит сложность сверхгибкой сутуры, а член lnk в правой части — константа. Изменчивость константы равна нулю, поэтому изменчивость сложности сверхгибкой сутуры должна равняться изменчивости логарифма радиуса раковины. Выбрав в качестве меры межвидовой изменчивости сигму, для сверхгибкой сутуры должны иметь σz = σlnR. <6.5> Таким образом, в пределе, когда требования к прочности раковины стали бы полностью удовлетворяться изменением длины сутуры, межвидовая изменчивость по ее сложности сравнялась бы с межвидовой изменчивостью по логарифму радиуса. Наши данные (рис. 30) показывают, что в ходе эволюции аммоноидей σz как бы стремится сравняться с σlnR, однако эта «цель» не достигается даже аммонитами юры и мела. По-видимому, какая-то часть требований среды к укреплению стенки раковины продолжала удовлетворяться за счет других средств. Тем не менее согласованное изменение сигмы обеих величин в течение последних 130 миллионов лет истории аммоноидей ярко демонстрирует, что технические возможности генетической системы, мобилизующей сутуру, фактически были доведены до предела. Прогресс в накоплении знаний Тезис «вид приспособлен к среде своего обитания» фактически банален, поскольку, раз мы имеем дело с видом, значит, он существует, а раз существует, значит, приспособлен. Можно сконструировать еще один тезис: «вид располагает знанием о среде своего обитания». Хотя это звучит несколько антропоморфно, формулировка не станет лучше, если мы заменим слово «знание» словом «информация». Можно различать два рода знаний вида о внешнем мире: знание видом собственной экологической ниши и знание им соседних ниш. Хотя утверждение о знаниях второго рода звучит несколько фантастично, однако это и есть то, что мы ранее назвали адаптируемостью. Знания первого рода воплощены в морфологии типичной особи, чьи рабочие структуры способны развивать мощность, достаточную для неопределенно долгого существования вида в одной и той же экологической нише. При сравнении гомологичных структур у разных видов можно заметить огромные различия в уровне их развития. Тем не менее, во всех случаях этот уровень (каким бы он нам ни представлялся), по-видимому, вид устраивает, а развивать орган сверх необходимости означало бы пустую трату средств, которые можно было бы направить на производство дополнительной биомассы. Природа как бы пытается снабдить организм рабочими структурами, способными развивать максимальную мощность при минимальных энергетических и информационных затратах. Заметим, что это чисто инженерная задача, и, как показывает наш практический опыт, при заданных материалах существует очень мало оптимальных решений (см. гл. 5). Знания второго рода касаются осведомленности вида о характере грядущих перемен, о преадаптированности к ним. Эти знания состоят в повышенной мобильности именно тех рабочих структур, от которых медленно изменяющаяся среда чаще всего требует усиления или ослабления. Мобильность структуры (как ответ на движущий отбор) обусловлена числом ее специальных генов и уровнем автономности развития ее зачатка. Число специальных генов определяет «выход» благоприятных мутаций, а уровень автономности — степень их благоприятности. Поэтому знания второй категории воплощены в сложности рабочих органов и в степени обособленности путей их индивидуального развития. Мобилизующий отбор ведет к накоплению филетической группой знаний прежде всего о своей адаптивной зоне, данный процесс не бесконечен и завершается установлением динамического равновесия между мобильностями рабочих структур. Это равновесие определяется характером изменчивости среды — частотой изменения ее требований к мощности той или иной структуры. Вполне понятно, что глобальные изменения катастрофического характера, уничтожая адаптивную зону, обесценивают и все знания о ней, накопленные филетической группой в ходе ее исторического развития. До сих пор остается открытым вопрос об эволюционных изменениях количества генетической информации. Простое измерение содержания ДНК в клеточных ядрах мало приближает нас к оценке числа генов, поскольку большую часть генома составляет ДНК межгенных промежутков и интронов. Не вдаваясь в анализ данных по многочисленным повторяющимся последовательностям, населяющим некодирующие районы генома, можно лишь отметить, что присутствие или отсутствие «повторов» очень мало сказывается на значениях основных компонентов приспособленности. Заметим, что у нас нет оснований считать, что отрезок ДНК содержит генетическую информацию, если его потеря не влияет на приспособленность организма. Мы уже отмечали, что для увеличения числа генов организмы должны искать пути к снижению мутационного генетического груза. Однако после разделения особей на два пола эволюция мало чего достигла на этом поприще. Эксперименты по насыщению деталями фиксированных отрезков генома позволяют оценить число генов, необходимых для жизни в «тепличных» условиях лаборатории. Такие оценки для мыши и дрозофилы оказались довольно близкими. Более того, 3–5 тысяч жизненно важных генов дрозофилы не намного больше 2 тысяч генов, которыми располагает «бесполая» кишечная палочка. Не исключено, что общее количество наследственной информации живых организмов вообще не изменялось в течение последнего миллиарда лет, т. е. с информационной точки зрения все живые организмы — и высшие и низшие — равны. Наш способ определения сложности на глаз ведет к сильной переоценке ее морфологической компоненты при явной недооценке химической. Достаточно вспомнить, что любая свободно живущая бактерия осуществляет в крошечном объеме своей клетки огромное число химических реакций, синтезируя с поразительной скоростью сложнейшие информационные макромолекулы. Как правило, бактерии не нуждаются в витаминах или незаменимых аминокислотах, а в качестве источников углерода, азота и фосфора могут использовать простые минеральные соли. Особенно впечатляет способность многих из них добывать энергию, используя такие вещества, как водород, сероводород, метан, сера, железо и т. д. Встает вопрос, почему именно многоклеточные вышли победителями в борьбе за место под солнцем? Что это так, свидетельствует хотя бы рис. 2, откуда мы видим, что по числу видов одноклеточные занимают в царстве животных одно из последних мест. По-видимому, главная причина этого явления заключается в том, что у многоклеточных относительно частые мутации, слегка изменяющие уровень генной активности, способны эффективно модулировать интенсивность основных функций организма. Это достигается созданием генетических программ развития надклеточных структур, когда работа более «ранних» генов вполне однозначно определяет время и место активации более «поздних». Такой характер межгенного взаимодействия означает, что мутация по любому из специальных генов (вне зависимости от их числа) может привести к изменению мощности рабочей структуры. В итоге даже небольшие изменения уровня генной экспрессии могут быть «замечены» и зафиксированы движущим отбором. Повышенная мобильность надклеточных структур, по-видимому, определяет макроэволюционную тенденцию к переходу генов из сферы клеточного домашнего хозяйства в сферу межклеточных взаимодействий. К тому же изменения экспрессии генов домашнего хозяйства могут повлиять на работу сразу всех клеток, что с определенной вероятностью ведет к нарушению координации основных функций организма, поэтому степень «благоприятности» соответствующих мутаций должна быть в среднем несколько ниже, чем мутаций по генам межклеточных взаимодействий с их более точным клеточным и функциональным адресом. Высокая эволюционная пластичность сложных надклеточных структур позволяет обладающим ими филетическим группам быстро проникать даже в другие адаптивные зоны (она как бы снабжает эти группы знанием о биосфере в целом). Достаточно отметить, с какой легкостью позвоночные после освоения суши завоевали воздух и вернулись (в новом качестве) в водную стихию. Теперь попробуем изложить в краткой форме суть нашей трактовки макроэволюционных тенденций. Суть нового подхода сводится к следующему. Пучок родственных филетических линий можно рассматривать как популяцию бесполых особей, чьи шансы на выживание определяются скоростью, с которой некоторые (лимитирующие) морфологические структуры организма способны изменять свою мощность для компенсации прогрессирующего ухудшения внешней среды. Иными словами, филетическое выживание видов определяется мобильностью лимитирующих структур, т. е. скоростью фиксации мутаций, изменяющих их мощность. Если каждая такая мутация изменяет мощность на сходный процент, то шансы вида на выживание будут тем выше, чем больше генов детерминирует развитие лимитирующей рабочей структуры. В то же время, при прочих равных условиях мощность морфологической структуры прямо связана с числом генов, детерминирующих ее развитие, поэтому чем мощнее будут лимитирующие структуры вида, тем выше его шансы на филетическое выживание. Таким образом, виды находятся под воздействием особой (мобилизующей) формы естественного отбора, направленного на повышение эволюционной пластичности организма, что скоррелировано с прогрессивным усложнением его морфологии. В итоге, все стороны строения особей можно рассматривать как результат длительного действия на их филетических предков отбора, повышающего адаптируемость видов к изменениям среды, наиболее типичным для адаптивной зоны. Существенный вклад в увеличение мобильности рабочей структуры вносит и повышенная скорость мутаций, изменяющих уровень экспрессии специальных генов. Независимость эффекта этих мутаций от числа специальных генов обеспечивается их подчинением особой программе развития, т. е. системе межгенных взаимодействий, управляющей индивидуальным развитием структуры. Такая организация специальных генов приводит к тому, что даже относительно небольшое изменение экспрессии любого из них ведет к заметному (для естественного отбора) сдвигу мощности соответствующей рабочей структуры. Очевидно также, что степень заметности этих мутаций для движущего отбора прямо связана с уровнем автономности программ развития разных рабочих структур. Таким образом, создается впечатление, что основные черты организации процесса индивидуального развития многоклеточных организмов сформировались под действием мобилизующего отбора.
Вместо заключения. Эволюционный прогресс: миф и реальность
У каждого века есть свои мифы. Их принято называть высшими истинами.Накопление филетической группой знаний о своей адаптивной зоне, естественно, увеличивает ее шансы на выживание. Постоянно идущий рост адаптируемости имеет многообщего с тем, что в отечественной биологической литературе традиционно называется эволюционным прогрессом. Например, известный советский эволюционист Л.Ш. Давиташвили предлагает такое определение: «Эволюционным прогрессом, или ароморфозом, мы будем называть такой тип эволюционного развития органических форм, который не только позволяет им существовать в занимаемой ими экологической обстановке, но и создает возможность непосредственного выхода этих форм или ближайших их потомков за пределы данной экологической обстановки». Однако что означает «выйти за пределы данной экологической обстановки»? Если речь идет о переходе в соседнюю экологическую нишу, а потом из нее в другую, подобную первой, то это одно; если же имеется в виду так называемый прорыв в новую адаптивную зону, то это совсем другое. В первом случае филетическая линия накапливает знания о своей адаптивной зоне, и этот процесс можно назвать прогрессом, во втором линия, безусловно, теряет знания о своей прошлой среде и приступает к накоплению знаний о совершенно новой. Таким образом, эволюционный прогресс может быть только относительным. Знания об адаптивной зоне виды воплощают в сложные полифункциональные рабочие структуры, способные быстро изменять свою мощность в ответ на наиболее типичные требования среды. Эти знания должны быть так или иначе записаны в нуклеотидных последовательностях ДНК, поэтому ограничения на объем генетической информации, накладываемые, прежде всего, мутационным генетическим грузом, ставят верхний предел возможностям эволюционирующей группы в овладении биосферой. Филетические линии постоянно «стремятся» как можно выше поднять уровень своей адаптируемости, однако этой тенденции противостоит принципиальная нестабильность среды, в частности, ограниченность во времени существования любой адаптивной зоны. Естественно, исчезновение адаптивной зоны обесценивает все завоевания предыдущей эволюции. Технический прогресс, безусловно, имеет много общего с эволюционным. Человечество, как и эволюционирующий филетический пучок, постоянно накапливает знания об окружающем мире и воплощает их в свои рабочие структуры (машины и механизмы), направляющие поток энергии и вещества из внешнего мира к людской биомассе. В обоих случаях наблюдается стремление к экономии энергии и материалов. Человечеству пока удается стремительно увеличивать объем знаний. Оглядываясь на свою историю, мы видим, что как-то умудрялись удваивать потребление энергии каждые 20 лет. Об экспоненциальном росте научно-технической литературы и упоминать нечего. Следовательно, наши знания о мире и технические возможности продолжают стремительно расти, не обнаруживая никаких признаков замедления. Естественно, такое положение вещей породило представление о безграничности процесса познания мира, а стало быть, и о безграничности наших технических возможностей. Это широко распространенное мнение остается основой для одного из мифов XIX–XX веков — мифа о техническом прогрессе. Перенесение этой, по существу, антропоморфной идеи на органический мир породило представление о безграничном эволюционном прогрессе, важнейшей чертой которого является увеличение потребления организмом энергии и возрастающая независимость его от среды. Нам кажется, что мы, стремительно увеличивая свои знания о среде с помощью нашего интеллекта и библиотек, являем собой воплощение древней эволюционной тенденции. Правда, кривые роста сложности и мощности рабочих структур совсем не похожи на устремленные в бесконечность экспоненты. Всюду мы видим, как такие кривые выходят на плато, ярко демонстрируя относительный и ограниченный характер эволюционного прогресса. Что же касается представления о нашем безграничном техническом прогрессе, то пока мы живем в эпоху экспоненциального роста потребления человечеством энергии и вещества, хотя ниоткуда не следует, что такое положение вещей будет сохраняться вечно. Во-первых, запасы энергии на Земле не безграничны, а, во-вторых, за нарастающей кривой потребления энергии и вещества как тень следует точно такая же кривая роста наших отбросов. Эта простая связь между техническим прогрессом и загрязнением среды все четче осознается. И уже слышны возгласы: «Хватит!» Еще одно ограничение на технический прогресс должно быть связано с небесконечными возможностями нашего интеллекта. Объем знаний, которым реально обладает и пользуется средний человек, едва ли возрастает. Мы учимся примерно столько же и примерно по той же методике, как учились наши предки в XVIII–XIX веках. Прогрессивное увеличение знаний обеспечивается лишь за счет стремительно сужающейся специализации. Ярким свидетельством тому является тезис: «Время энциклопедистов прошло». Однако уже появились первые предвестники грядущего ослабления информационного потока, затопляющего технические отделы наших библиотек. Все больше начинают цениться гуманитарные знания, которые не повышают приток энергии к нашей биомассе, а служат удовлетворению духовных запросов. Похоже на то, что время технократов идет к концу, а с ним завершится и фаза нашего безудержного технического прогресса. В создании мифа об эволюционном прогрессе немалую роль сыграла безотчетная склонность людей объяснять природные явления с помощью законов, совершенно жестко, т. е. с абсолютной необходимостью связывающих наблюдаемые события. Такие динамические законы позволяют (при наличии исчерпывающей информации) со 100 %-й вероятностью предсказывать будущее состояние системы любой сложности. В отношении живых систем данный подход ведет к признанию особых, пока не открытых сил, толкающих живую материю в сторону повышения организации. Эта простая, а потому и привлекательная идея, впервые выдвинутая Ж.-Б. Ламарком, продолжает и поныне сохранять немалое число приверженцев. Психологический момент, затрудняющий восприятие случайности как фундаментального свойства материи, ярко отразился в негативном отношении А. Эйнштейна к статистическим основам квантовой физики. В письме Дж. Франку он писал: «Я еще могу представить, что бог создал мир, в котором нет законов природы, короче говоря, что он создал хаос. Но чтобы статистические законы были окончательными, и бог разыгрывал каждый случай в отдельности — такая мысль мне крайне несимпатична». И тем не менее, фундаментальный характер случайности некоторых явлений, имеющих принципиальное значение для эволюционного процесса, не вызывает сомнений. Наиболее важным здесь представляется отсутствие какой-либо корреляции между величиной и знаком мутационного эффекта на функцию структуры, с одной стороны, и сдвигом среды, изменяющим ее требование к этой структуре, — с другой. В данной книге мы попытались показать, как статистические по своему характеру закономерности способны дать удовлетворительное объяснение феномену прогрессивной эволюции без привлечения каких-либо особых, неизвестных биологам динамических факторов.Неизвестный автор[1]
Последние комментарии
1 день 6 часов назад
1 день 8 часов назад
1 день 23 часов назад
1 день 23 часов назад
2 дней 4 часов назад
2 дней 8 часов назад