Дэвид Хантер Хьюбел Глаз, мозг, зрение
Предисловие к русскому изданию
Если попросить исследователей зрительной системы, придерживающихся самых различных точек зрения на функцию нейронов зрительной коры, назвать одно наиболее яркое их свойство, без сомнения, будет названа ориентационная избирательность. Она состоит в том, что отдельные клетки реагируют только на линии определенного направления (например, горизонтальные), появившиеся в поле зрения животного. Открытие такой избирательности, сделанное в 1961 году автором этой книги совместно с Т. Визелом, постоянным его соавтором на протяжении последующих двадцати лет, положило начало современному этапу в изучении организации зрительной коры высших млекопитающих. За открытием ориентационной избирательности последовали и другие фундаментальные результаты: первая классификация нейронов зрительной коры, выявление групп нейронов («колонок»), сходных по функциональным свойствам, работы, показавшие значение раннего полноценного зрительного опыта для нормального созревания и развития генетически предопределенных структур зрительной коры, и другие исследования. Д. Хьюбел и Т. Визел относятся к числу исследователей, имеющих относительно небольшое число публикаций, но почти каждая их работа была новой главой в физиологии центральных отделов зрительной системы. Этим авторам не свойственно широкое теоретизирование и построение фундаментальных, но не привязанных к экспериментальной базе концепций о функционировании мозга или зрительной коры. В одной из своих работ они говорили, что, к счастью, спекуляции о принципах работы мозга не являются единственным путем для исследования и что изучать мозг — более увлекательное и, кажется, более полезное занятие. Каждая их работа имела простую и четкую задачу, для решения которой ставился адекватный, обычно несложный эксперимент, совсем не обязательно использующий все последние технические возможности. В то же время Д. Хьюбел был одним из первых, кто понял и применил на практике новые гистохимические методики для морфологического выявления ориентационных и глазодоминантных колонок. Особенно изящными были эксперименты, выполненные (совместно с М. Ливингстон) уже после присуждения ему и Т. Визелу в 1981 году Нобелевской премии. Эти работы, сочетающие электрофизиологический эксперимент с изучением локальных связей в коре методом микроинъекций пероксидазы хрена, привели к открытию мозаичной структуры зрительных зон коры. И, как часто бывало и раньше, эта особенность организации зрительных зон оказалась общим принципом организации и других отделов коры мозга. Все сказанное нашло яркое отражение в книге Д. Хьюбела. Он как бы приглашает проследить за историей развития своих исследований и ведет от проблемы к проблеме. Стараясь быть понятным широкому кругу читателей, в том числе не имеющих запаса физиологических знаний, автор часто обращается к их здравому смыслу. В книге не только описаны оригинальные исследования автора, но и изложены его представления о работе всей зрительной системы. Раздел, посвященный глазу и сетчатке, по необходимости содержит довольно много чисто учебного материала, но не может претендовать на полноту и адекватность описания. К примеру, ни словом не упомянуто о такой удивительной особенности организации сетчатки, как широкая электрическая связь между фоторецепторами и особенно между горизонтальными клетками, о «детекторной» концепции Дж. Леттвина с соавторами, разработанной в 1959 году и сыгравшей весьма конструктивную роль в последующих исследованиях сетчатки и зрительной системы, особенно у низших позвоночных. С другой стороны, в книге хорошо представлены сведения о цветовом зрении человека, включая такое важное его свойство, как константность цветовосприятия, и о бинокулярном зрении. Можно не сомневаться, что книга Д. Хьюбела вызовет большой интерес у широкого круга читателей, не только нейрофизиологов, офтальмологов и просто любопытных, но и у всех, кто занимается такими ныне популярными областями исследований, как нейронные сети и искусственный интеллект.А. Л. Бызов И. Н. Пигарев
Предисловие
Эта книга посвящена в основном развитию наших представлений о том, каким образом мозг обрабатывает зрительную информацию; она охватывает период примерно с 1950 по 1980 г. и касается главным образом исследований, в которых я либо участвовал сам, либо был в значительной мере заинтересован. Мне, я считаю, повезло работать в эпоху, которую можно назвать волнующей и радостной. Некоторые эксперименты были невероятно трудными — во всяком случае, они казались такими в пятом часу утра, особенно если опыт проходил неудачно. В целом, однако, работа доставляла наслаждение. В нейрофизиологических экспериментах есть особая непосредственность: реакцию клетки на стимуляцию можно видеть и слышать, и нередко сразу удается сообразить, какова роль таких реакций в функционировании мозга. Нейробиология занимает особое положение в современной науке еще и по мизерности (если сравнивать с ядерной физикой или астрономией) того бюджета, в рамках которого вы можете работать в одиночку или с единственным помощником. Особой удачей было получить образование и работать на североамериканском континенте, где замечательная университетская система сочетается с правительством, постоянно финансирующим исследования по биологии, особенно в области зрения. Я могу лишь надеяться, что у нас хватит здравого смысла, чтобы бережно сохранять подобные преимущества. При написании этой книги я имел в виду такого читателя, как, например, астроном, т.е. человека с научным образованием, но не специалиста по биологии и тем более по нейробиологии. Я старался дать ровно столько фундаментальных сведений, сколько нужно, чтобы сделать понятными нейробиологические проблемы, не перегружая текст материалом, представляющим интерес лишь для специалистов. Проложить курс между чрезмерной поверхностностью и излишней детализацией оказалось не просто, особенно если учесть саму природу мозга, заставляющую нас выводить из множества четких взаимосвязанных деталей какое-то представление о том, что собой представляет объект изучения и что он делает. Все описанные здесь исследования, в которых я принимал участие, явились результатом совместных усилий. С 1958 до конца 70-х годов моим партнером по работе был Торстен Визел. Если бы не его идеи, энергия, энтузиазм, выдержка и старание ужиться с раздраженным коллегой, каким был я, результат был бы совершенно иным. Мы оба бесконечно признательны Стивену Куффлеру, который на ранних этапах весьма тактично направлял нашу работу, поощрял нас своим беспредельным энтузиазмом, а иногда и развенчивал наши тщетные претензии попросту тем, что выглядел озадаченным. Нейробиологическая группа отделения фармакологии Гарвардской медицинской школы (1963). Из этой группы позднее было образовано Отделение нейробиологии. Стоят (слева направо): Эдвин Фершпан, Стивен Куффлер, Дэвид Хьюбел; сидят (слева направо): Дэвид Поттер, Эдуард Кравитц, Торстен Визел.
Нейробиологическая группа отделения фармакологии Гарвардской медицинской школы (1963). Из этой группы позднее было образовано Отделение нейробиологии. Стоят (слева направо): Эдвин Фершпан, Стивен Куффлер, Дэвид Хьюбел; сидят (слева направо): Дэвид Поттер, Эдуард Кравитц, Торстен Визел.
При написании книги необходима критика (мне во всяком случае), и чем суровей и беспощадней, тем лучше. Я особенно признателен Эрику Кэнделу за его помощь в расстановке акцентов в первых трех главах, а также моей сотруднице Маргарет Ливингстон, разбившей буквально в пух и прах содержание еще трех глав. Одно из ее замечаний начиналось так: «Сначала Вы напускаете туману, а потом просто пишете не то...» Ей приходилось быть терпимой к моей вспыльчивости и мириться с отсрочкой собственных исследований. Я также признателен редакторам Научной американской библиотеки, особенно Сьюзан Моран, Линде Дэвис, Джерарду Пилу и Линде Чапут, а также корректору Синтии Фарден: я не мог ранее вообразить, насколько качество текста будет зависеть от умелых и увлеченных редакторов. Они исправили бесчисленные грамматические ошибки, но помощь их простиралась значительно дальше этого: они выявляли повторы, улучшали ясность изложения и были терпимыми к моей склонности ставить запятые там, где не нужно. Кроме всего прочего, они не переставали вгрызаться в меня до тех пор, пока я не излагал свои мысли в легко доступной (как я надеюсь!) форме. Я хочу поблагодарить Кэрол Доннер за художественное оформление, а также дизайнера Нэнси Филд, координатора по иллюстрациям Мелани Нильсен и производственного координатора Сьюзан Стетцер. Я также благодарю за критическое прочтение книги Сьюзан Эбукайр, Дэвида Кардосо, Уитмора Тингли, Дебору Гордон, Ричарда Мэсланда и Лауру Риген. Как всегда, незаменимой оказалась моя секретарша Оливия Брам, весьма терпеливая к моему нраву. Жена Рут помогала мне многочисленными советами, а также тем, что примирилась со множеством потерянных уикендов. Теперь будет большим облегчением не слышать, как мои дети говорят: «Папочка, ну когда же ты, наконец, закончишь эту свою книгу?» Временами этот день казался чем-то столь же отдаленным, как остров Санчо Пансы.
Дэвид Хьюбел
1. Введение
Наш мозг сложен — говорит интуиция. Мы осуществляем огромное множество сложных актов. Мы дышим, кашляем, чихаем, совокупляемся, глотаем, извергаем содержимое желудка обратно, мочимся; мы складываем и вычитаем, разговариваем и даже рассуждаем, пишем, поем и сочиняем квартеты, стихи, романы и пьесы; мы играем в бейсбол и на музыкальных инструментах. Мы воспринимаем и думаем. Может ли орган, управляющий всеми этими действиями, не быть сложным? Следует ожидать, что орган со столь богатыми возможностями должен иметь очень сложное строение. Можно думать, что он по крайней мере состоит из большого числа элементов. Одно это, однако, еще не гарантирует его сложности. Головной мозг содержит 1012 (миллион миллионов) клеток — по любым понятиям число астрономическое; хотя я не знаю, подсчитывал ли кто-нибудь число клеток в печени человека, было бы удивительно, если бы в ней было меньше клеток, чем в нашем мозгу. И все же никто никогда не скажет, что печень так же сложна, как мозг. Более веский аргумент в пользу сложности мозга можно усмотреть в соединениях между его клетками. Обычная нервная клетка (нейрон) получает здесь информацию от сотен или тысяч других клеток и в свою очередь передает информацию сотням или тысячам нейронов. Общее число соединений в мозгу, таким образом, должно составлять приблизительно 1014–1015. Но как это число ни огромно, все же само по себе оно еще не служит надежным показателем сложности. Анатомическая сложность определяется не только числом элементов, но и характером организации, который трудно оценить числами. Можно проводить аналогии между мозгом человека и гигантским органом, печатной машиной, телефонной станцией или большим компьютером, но польза от подобных аналогий заключается главным образом в наглядном представлении о множестве малых частей, смонтированных в определенном точном порядке, функции которых по отдельности или вместе неспециалист уловить не может. В сущности, такие аналогии полезнее для тех, кто совсем не имеет понятия, как работают печатные машины и телефонные станции. В конце концов, для того чтобы получить представление о том, что такое мозг, как он устроен и как обрабатывает информацию, нет другого пути, кроме детального ознакомления с самим мозгом или его частями. В этой книге я надеюсь разъяснить читателю некоторые особенности структуры и функции мозга, подробно рассмотрев ту его часть, с которой связано зрение. Вопросы, которых я буду касаться, можно сформулировать достаточно просто. Когда мы смотрим на окружающий мир, первичное событие состоит в фокусировке света на сетчатке каждого глаза. Сетчатка содержит 125 миллионов рецепторов, называемых палочками и колбочками; это нервные клетки, специализированные таким образом, чтобы генерировать электрические сигналы при попадании на них света. Задача остальной части сетчатки и самого мозга — использовать эти сигналы, чтобы извлечь биологически полезную информацию. Результатом будет зрительная сцена в том виде, как мы ее воспринимаем, со всей сложностью форм, глубины, движения, цвета и текстуры. Мы хотим узнать, каким образом мозг решает эту сложнейшую задачу. Рис. 1. Сантьяго Рамон-и-Кахал играет в шахматы в 1898 году в возрасте 46 лет во время отпуска в Мирафлорес де ла Сьерра. Снимок сделан одним из его детей. По мнению большинства нейроанатомов, Рамон-и-Кахал намного превзошел любого специалиста в этой области, а может быть, и во всей нейробиологии центральной нервной системы. Его двумя главными достижениями явились: 1) весьма убедительная демонстрация того, что нервные клетки действуют как независимые единицы, и 2) использование метода Гольджи для картирования обширных участков головного и спинного мозга, показавшее как чрезвычайную сложность, так и высокую упорядоченность нервной системы. В 1906 году ему вместе с Гольджи была присуждена Нобелевская премия.
Рис. 1. Сантьяго Рамон-и-Кахал играет в шахматы в 1898 году в возрасте 46 лет во время отпуска в Мирафлорес де ла Сьерра. Снимок сделан одним из его детей. По мнению большинства нейроанатомов, Рамон-и-Кахал намного превзошел любого специалиста в этой области, а может быть, и во всей нейробиологии центральной нервной системы. Его двумя главными достижениями явились: 1) весьма убедительная демонстрация того, что нервные клетки действуют как независимые единицы, и 2) использование метода Гольджи для картирования обширных участков головного и спинного мозга, показавшее как чрезвычайную сложность, так и высокую упорядоченность нервной системы. В 1906 году ему вместе с Гольджи была присуждена Нобелевская премия.
Чтобы ваши надежды и ожидания не были чрезмерными, я должен буду вас предупредить, что мы знаем лишь небольшую часть ответа. Однако нам уже многое известно о механизмах зрительной системы, и мы имеем довольно ясное представление о том, как мозг приступает к решению этой задачи. Наши познания достаточно убеждают в том, что мозг, хотя и сложным образом, работает в соответствии с принципами, которые когда-то, вероятно, будут поняты, и что ответы будут не столь сложны, чтобы их можно было понять лишь обладателям ученых степеней в области компьютерной науки или физики элементарных частиц. Нейробиология — увлекательный, но необычный предмет. Она имеет дело со структурой нервной системы (нейроанатомией) и с ее функцией (нейрофизиологией). Биология большинства систем органов рассматривает форму, относительное расположение и работу таких объектов, как кости, пищеварительный тракт, почки или печень, функции которых сравнительно хорошо известны. Я не утверждаю, что о каждом из них известно абсолютно все, но у нас есть хотя бы приблизительные представления: пищеварительный тракт перерабатывает пищу, сердце перекачивает кровь, кости обеспечивают опору, а в некоторых костях происходит образование крови. (Сейчас трудно представить себе эпоху, будь то даже темное двенадцатое столетие, когда люди не понимали бы, что благодаря костям человек отличается по консистенции от дождевого червя; мы, однако, легко забываем, что для открытия роли сердца понадобился уже гений Уильяма Гарвея.) Для чего служит нечто? — это вопрос, применимый только к объектам живой природы (и ее производным). Можно спросить, и этот вопрос осмыслен, для чего нужны ребра: они образуют грудную клетку и препятствуют сдавлению находящихся в ней органов. Можно также спросить, для чего служит мост: люди, будучи частью живой природы, изобрели его, чтобы переправляться через реку. Вне биологии цель смысла не имеет, именно поэтому я улыбаюсь, услышав вопрос сына: «Папа, зачем идет снег?». Возникновение цели в живой природе связано с эволюцией, борьбой за существование, социобиологией, генами самосохранения — множеством высоких материй, постоянно занимающих умы многих людей. Спустимся, однако, на землю. Большинству анатомических объектов, даже таким некогда загадочным образованиям, как зобная железа и селезенка, ныне можно приписать вполне осмысленные функции. Когда я был еще студентом-медиком, слова о зобной железе и селезенке сопровождали вопросительными знаками. Иначе обстоит дело с мозгом, ибо даже сейчас обширные его участки помечают вопросительными знаками, размышляя не только о том, как они работают, но и об их биологическом значении. Огромная, детально разработанная область нейроанатомии — это в значительной части нечто вроде географии структур, функции которых все еще остаются загадкой или не вполне ясны. Степень нашего невежества, конечно, не одинакова. Например, мы знаем довольно много о зоне, называемой двигательной корой, и имеем общее представление об ее функции: она обслуживает произвольные движения; стоит только разрушить ее на одной стороне, как рука и нога на противоположной стороне станут неуклюжими и слабыми и мимика половины лица будет нарушена. Уровень наших знаний о двигательной коре находится где-то посередине всей шкалы от абсолютного незнания функций некоторых структур мозга до ясного понимания немногих других его структур (подобно тому как большинство из нас не имеет понятия о работе компьютера, лучше понимает работу печатного станка, еще лучше — двигателя внутреннего сгорания и совсем хорошо — устройства, которое мы сами изобрели). Зрительный путь, включая первичную зрительную кору (стриарную кору), относительно хорошо исследован. Зрительная кора — быть может, лучше всего изученная ныне часть мозга и уж наверняка наиболее известная часть коры его больших полушарий. Мы довольно хорошо знаем, «для чего» она нужна, т.е. чем обычно заняты ее нервные клетки, и приблизительно представляем себе ее вклад в анализ зрительной информации. Такого знания мы достигли сравнительно недавно, и я хорошо помню, как в 1950-е годы я рассматривал под микроскопом срез зрительной коры с бесчисленными клетками, упакованными как яйца в коробке, и размышлял о том, что́ бы они все вместе могли делать и сможем ли мы когда-нибудь это узнать.
 Рис. 2. Головной мозг человека — вид слева и несколько сзади; можно видеть кору больших полушарий и мозжечок. Непосредственно перед мозжечком виден небольшой участок ствола.
Рис. 2. Головной мозг человека — вид слева и несколько сзади; можно видеть кору больших полушарий и мозжечок. Непосредственно перед мозжечком виден небольшой участок ствола.
Каким образом нужно было бы приступать к выяснению этого? Первая пришедшая в голову мысль могла состоять в том, что подробное рассмотрение связей между глазом и мозгом и внутри мозга окажется уже достаточным, чтобы заключить о том, как они работают. К сожалению, это верно лишь отчасти. Давно было известно, что для зрения важны области коры, расположенные в задней части нашего мозга, — еще на рубеже нынешнего века было обнаружено, что глаза связаны с нею через промежуточную «узловую станцию». Но чтобы понять, основываясь только на структурных данных, что делают клетки зрительной коры, когда животное или человек смотрит на небо или на дерево, необходимо знать анатомические детали намного лучше, чем мы знаем их сейчас. Нелегко нам было бы даже в том случае, если бы мы имели полную схему всех связей, точно так же как трудно понять устройство компьютера или радиолокационной установки только по их схемам, особенно если мы не знаем, для чего предназначены эти устройства. Прогресс в понимании работы зрительной коры явился результатом применения комбинированной стратегии. Еще в конце 50-х годов физиологический метод регистрации активности одиночных нейронов начал доставлять кое-какие сведения о том, как функционируют нейроны в повседневной жизни животного; между тем существенных успехов в построении детальной схемы связей в то время не было. Однако в последние несколько десятилетий обе области — физиология и анатомия — параллельно продвигались вперед, взаимно обогащая друг друга методиками и новой информацией. Иногда говорят, что нервная система содержит огромное число случайных межнейронных соединений. Хотя упорядоченность связей и в самом деле не всегда очевидна, я подозреваю, что те, кто говорит о случайных нейронных сетях, не утруждали себя ознакомлением с нейроанатомией. Даже беглый просмотр такой книги, как труд Кахала «Histologie du Systéme Nerveux», достаточно убеждает кого угодно в том, что в чудовищной сложности нервной системы почти всегда можно усмотреть известную степень упорядоченности. Когда мы видим правильные ряды клеток в мозгу, впечатление создается такое же, как если бы мы рассматривали телефонную станцию, печатный станок или внутренность телевизора, — становится несомненным, что упорядоченность служит какой-то цели. Столкнувшись с тем или иным человеческим изобретением, мы едва ли усомнимся в том, что агрегат в целом, так же как и его отдельные части, обладает вполне постижимыми функциями. Чтобы понять их, нам нужно только прочесть ряд инструкций. В биологии появляется аналогичная вера в функциональную обоснованность и в конечном итоге даже в постижимость структур, которые не были изобретены кем-то, а совершенствовались на протяжении миллионов лет эволюции. Задача нейробиолога (разумеется, не единственная) состоит в том, чтобы выяснить связь порядка и сложности с функцией. Для начала я хочу дать упрощенное представление о том, на что похожа нервная система — как она построена, как работает и как мы собираемся изучать ее. Я опишу типичные нервные клетки и структуры, которые из них создаются. Основные «строительные блоки» в мозгу — нейроны, или нервные клетки. Это не единственные клетки в нервной системе: в перечень структурных элементов мозга следует также включить глиальные клетки, которые скрепляют нейроны и, вероятно, помогают питать их и удалять ненужные продукты обмена веществ; кровеносные сосуды и составляющие их клетки; различные покрывающие мозг оболочки; и даже, пожалуй, череп, который вмещает остальные структуры и обеспечивает их защиту. Здесь я буду рассматривать только нервные клетки. Многие видят в нервах подобие нитевидных проводов, по которым распространяются электрические сигналы. Но нервное волокно — это только одна из многих частей нейрона. Тело нейрона имеет обычно более или менее шаровидную форму, свойственную многим клеткам (см. рис. 3), и содержит ядро, митохондрии и другие органеллы, выполняющие многочисленные «внутрихозяйственные» функции, о которых так любят говорить цитофизиологи. От тела клетки отходит главный отросток в виде цилиндрической нити — нервное волокно, передающее сигнал и называемое аксоном. Кроме аксона от тела отходит множество других ветвящихся и суживающихся к концу волокон; их называют дендритами. Вся нервная клетка — ее тело, аксон и дендриты — одета клеточной мембраной. Тело нейрона и дендриты получают информацию от других нейронов; аксон передает информацию от данного нейрона другим нейронам. Длина аксона варьирует в пределах от долей миллиметра до метра и более; длина большинства дендритов не превышает миллиметра. Вблизи своего окончания аксон обычно разделяется на многочисленные веточки, концевые участки которых очень близко подходят к телам или дендритам других нервных клеток, но не соприкасаются с ними вплотную. В этих областях, называемых синапсами, информация передается от одной нервной клетки, пресинаптической, к следующей — постсинаптической.
 Рис. 3. Главные части нервной клетки — это ее тело, содержащее ядро и другие органеллы, единственный аксон, передающий импульсы от клетки, и дендриты, к которым приходят импульсы от других клеток.
Рис. 3. Главные части нервной клетки — это ее тело, содержащее ядро и другие органеллы, единственный аксон, передающий импульсы от клетки, и дендриты, к которым приходят импульсы от других клеток.
Сигналы в нерве возникают в точке аксона, близкой к месту его соединения с телом клетки; они передаются вдоль аксона, удаляясь от тела клетки, и доходят до области концевых разветвлений. Из окончаний аксона информация передается через синапсы следующей клетке или клеткам — здесь происходит химическая передача, которую мы рассмотрим в главе 2. Нервные клетки далеко не одинаковы, они делятся на множество различных типов. Хотя есть и промежуточные формы, в целом это деление на типы достаточно четко. Никто не знает, сколько типов существует в головном мозгу, — их, несомненно, больше сотни, а может быть, и больше тысячи. Нет двух совершенно одинаковых нейронов. Две клетки одного и того же класса примерно так же сходны между собой, как два дуба или два клена, а различие между двумя классами можно сравнить с отличием кленов от дубов или даже от одуванчиков. Не следует рассматривать классы клеток как жесткие подразделения: в зависимости от вашей склонности к дроблению или к объединению вы, возможно, насчитаете в сетчатке и в коре мозга по полсотне типов клеток или всего лишь по полудюжине типов (см. примеры на рис. 4).
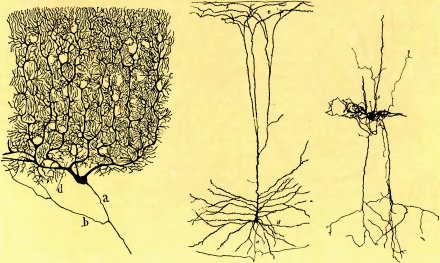 Рис. 4. Слева: мозжечковая клетка Пуркинье, зарисованная Сантьяго Рамон-и-Кахалом. Это один из крайних случаев специализации нейрона. Густое древовидное ветвление дендритов по форме напоминает не куст, а ветвь кедра, так как все разветвления расположены в одной плоскости. Через незаполненные веточками участки, напоминающие отверстия, проходят миллионы тончайших аксонов, идущих наподобие телеграфных проводов под прямым углом к плоскости рисунка. Аксон клетки Пуркинье отдает несколько веточек поблизости от тела клетки, а затем спускается к клеточным скоплениям, расположенным в глубине мозжечка на расстоянии нескольких сантиметров, где он расщепляется на многочисленные концевые разветвления. Полная высота клетки (тело плюс дендриты) в реальном масштабе составляет около 1 миллиметра.
В середине: сделанная Рамон-и-Кахалом зарисовка пирамидного нейрона коры больших полушарий, окрашенного по методу Гольджи. Общая высота рисунка соответствует примерно 1 миллиметру. Показана лишь часть (а) главного аксона: отдав две веточки (с), он может продолжаться за пределы рисунка на расстояние нескольких сантиметров (и даже метров), прежде чем окончится густой сетью разветвлений. Небольшое черное пятнышко — тело нейрона.
Справа: на этом рисунке Йеннифера Лунда изображена корковая клетка, которую относят к типу звездчатых нейронов. Темный пузырек в центре — тело клетки. Аксоны (тонкие линии) и дендриты (толстые линии) ветвятся и идут вверх и вниз на расстояние около миллиметра.
Рис. 4. Слева: мозжечковая клетка Пуркинье, зарисованная Сантьяго Рамон-и-Кахалом. Это один из крайних случаев специализации нейрона. Густое древовидное ветвление дендритов по форме напоминает не куст, а ветвь кедра, так как все разветвления расположены в одной плоскости. Через незаполненные веточками участки, напоминающие отверстия, проходят миллионы тончайших аксонов, идущих наподобие телеграфных проводов под прямым углом к плоскости рисунка. Аксон клетки Пуркинье отдает несколько веточек поблизости от тела клетки, а затем спускается к клеточным скоплениям, расположенным в глубине мозжечка на расстоянии нескольких сантиметров, где он расщепляется на многочисленные концевые разветвления. Полная высота клетки (тело плюс дендриты) в реальном масштабе составляет около 1 миллиметра.
В середине: сделанная Рамон-и-Кахалом зарисовка пирамидного нейрона коры больших полушарий, окрашенного по методу Гольджи. Общая высота рисунка соответствует примерно 1 миллиметру. Показана лишь часть (а) главного аксона: отдав две веточки (с), он может продолжаться за пределы рисунка на расстояние нескольких сантиметров (и даже метров), прежде чем окончится густой сетью разветвлений. Небольшое черное пятнышко — тело нейрона.
Справа: на этом рисунке Йеннифера Лунда изображена корковая клетка, которую относят к типу звездчатых нейронов. Темный пузырек в центре — тело клетки. Аксоны (тонкие линии) и дендриты (толстые линии) ветвятся и идут вверх и вниз на расстояние около миллиметра.
Связи между нейронами или группами нейронов мозга обычно не очевидны, и для того чтобы выявить наиболее важные проводящие пути, потребовались столетия. Поскольку в густых сетях волокон разные их пучки часто пронизывают друг друга, для изучения каждого пучка в отдельности нужны специальные методы. Выбранный для исследования участок мозга может быть невероятно плотно заполнен клеточными телами, дендритами и аксонами, между которыми почти нет свободного пространства. Поэтому методы окрашивания клеток, способные выявить и раскрыть организацию более рыхлых клеточных структур, таких как печень или почка, дают в мозгу лишь сплошную черную массу. Нейроанатомы, однако, придумали новые эффективные способы выявления как отдельных клеток в какой-либо одиночной структуре, так и связей между разными структурами. Как можно было ожидать, нейроны с одинаковыми или близкими функциями часто бывают связаны между собой. Тесно взаимосвязанные клетки в нервной системе нередко группируются вместе по очевидной причине большей эффективности коротких аксонов: такие аксоны «дешевле изготовить», они занимают меньше места и быстрее доставляют сообщения своим адресатам. Мозг поэтому содержит сотни клеточных скоплений, которые могут иметь форму шаров или параллельных слоев. Кора большого мозга — пример такой одиночной гигантской пластины толщиной два миллиметра и площадью около квадратного фута. В отдельном скоплении между нейронами могут быть короткие связи, а иногда от одного скопления к другому идет большое число длинных волокон, образующих пучки, или тракты. Шаровидные или пластинчатые структуры часто соединяются последовательно в проводящие пути (см. рис. 6).
 Рис. 5. На этом рисунке Рамон-и-Кахала (препарат, окрашенный по Гольджи) видны несколько клеток в верхних слоях коры мозга месячного младенца. В этой области окрасилась лишь ничтожная доля клеток.
Рис. 5. На этом рисунке Рамон-и-Кахала (препарат, окрашенный по Гольджи) видны несколько клеток в верхних слоях коры мозга месячного младенца. В этой области окрасилась лишь ничтожная доля клеток.
 Рис. 6. Зрительный путь. Каждая структура, представленная квадратиком, содержит миллионы клеток, расположенных слоями, имеет входы от одной или нескольких структур нижележащих уровней и посылает выходные сигналы нескольким структурам вышележащих уровней. Вверх от первичной зрительной коры этот путь прослежен на протяжении всего лишь четырех или пяти уровней.
Рис. 6. Зрительный путь. Каждая структура, представленная квадратиком, содержит миллионы клеток, расположенных слоями, имеет входы от одной или нескольких структур нижележащих уровней и посылает выходные сигналы нескольким структурам вышележащих уровней. Вверх от первичной зрительной коры этот путь прослежен на протяжении всего лишь четырех или пяти уровней.
Хорошим примером такой цепи последовательных структур служит зрительный путь. Сетчатка каждого глаза состоит из трех слоев клеток, один из которых содержит светочувствительные рецепторные клетки — палочки и колбочки. Как уже говорилось, в каждом глазу имеется более 125 миллионов рецепторов. Две сетчатки посылают свои выходные сигналы двум клеточным скоплениям величиной с ядро арахиса, расположенным в глубине мозга, — так называемым наружным (латеральным) коленчатым телам. От этих структур в свою очередь идут волокна в зрительную область коры. Точнее, они направляются к стриарной коре — первичной зрительной зоне. Отсюда, пройдя через ряд слоев синаптически связанных клеток, информация передается соседним зрительным зонам более высокого порядка, а те посылают сигналы нескольким другим зонам (см. рис. 6). Каждая из этих корковых зон, подобно сетчатке, содержит три или четыре синаптических переключения. В самом заднем отделе коры — затылочной доле — имеется не меньше дюжины таких зрительных зон (каждая величиной примерно с почтовую марку), и намного большее их число, видимо, располагается в теменной и височной долях, лежащих перед затылочной. Здесь, однако, наши знания о зрительном пути становятся смутными. Наша основная цель состоит сейчас в том, чтобы понять, для чего существуют все эти цепочки нейронных структур, как они работают и что делают. Мы хотим узнать, какого рода зрительная информация передается по описанным путям и как она модифицируется на каждом этапе — в сетчатке, наружном коленчатом теле и на различных уровнях коры. Мы изучаем эту проблему с помощью микроэлектрода — важнейшего инструмента современной эры в нейрофизиологии. Мы вводим микроэлектрод (обычно тонкую изолированную проволочку) в ту структуру, которую хотим исследовать, — например в наружное коленчатое тело, — так, чтобы его кончик, подойдя к клетке достаточно близко, дал нам возможность регистрировать ее электрические сигналы. Мы пытаемся воздействовать на эти сигналы подачей различных световых стимулов на сетчатку животного.
 Рис. 7. Постановка эксперимента по регистрации ответов в зрительной системе. Животное (обычно это макак) помещают перед экраном, на который проецируется стимул. Регистрацию осуществляют, вводя микроэлектрод в тот или иной участок зрительного пути, в данном случае в первичную зрительную кору. (На рисунке изображен мозг человека, но мозг обезьяны очень сходен с ним.)
Рис. 7. Постановка эксперимента по регистрации ответов в зрительной системе. Животное (обычно это макак) помещают перед экраном, на который проецируется стимул. Регистрацию осуществляют, вводя микроэлектрод в тот или иной участок зрительного пути, в данном случае в первичную зрительную кору. (На рисунке изображен мозг человека, но мозг обезьяны очень сходен с ним.)
Наружное коленчатое тело получает входные сигналы главным образом от сетчатки — каждая его клетка имеет связи с палочками и колбочками, хотя и не прямые, а через промежуточные нейроны сетчатки. Как вы увидите в главе 3, палочки и колбочки, передающие сигналы той или иной клетке зрительного пути, не разбросаны по всей сетчатке, а собраны в небольшом участке. Этот участок называют рецептивным полем данной клетки. Поэтому наш первый шаг состоит в том, чтобы, освещая различные места сетчатки, найти это рецептивное поле. Определив его границы, мы можем начать варьировать форму, размеры, цвет и скорость перемещения светового стимула и таким образом выяснять, на какие виды зрительных стимулов лучшего всего реагирует изучаемая клетка. Нам нет надобности направлять пучок света прямо на сетчатку. Обычно гораздо проще проецировать стимулы на экран, расположив его в нескольких метрах от животного. Глаз создает на сетчатке хорошо сфокусированное изображение экрана и стимула. Теперь мы можем начать работу и определить на экране положение проекции рецептивного поля. Если угодно, можно рассматривать рецептивное поле как часть зрительного мира животного — в данном случае часть экрана, «видимую» интересующей нас клеткой. Мы вскоре узнаем, что реакции клеток могут быть — и обычно бывают — весьма избирательными. Для того чтобы подобрать стимул, вызывающий по-настоящему сильную реакцию данного нейрона, может потребоваться известное время на поиски. Сначала нам иной раз трудно будет даже найти место рецептивного поля на экране, хотя для клеток такого раннего этапа, как коленчатое тело, его можно легко локализовать. Клетки коленчатого тела избирательны как по отношению к величине пятна, на которое они реагируют, так и к тому, будет ли это черное пятно на белом фоне или белое на черном фоне. На более высоких уровнях мозга для вызова реакции у некоторых клеток может потребоваться стимул в виде «края» (границы света и темноты); при этом часто будет иметь значение ориентация края — расположен ли он вертикально, горизонтально или наклонно. Иногда бывает важно, неподвижен стимул или он движется по сетчатке (или экрану), цветной он или белый. Если на экран смотрят оба глаза, решающим может быть точное расстояние до экрана. Отдельные нейроны, даже в пределах одной и той же структуры, могут сильно различаться по стимулам, на которые они реагируют. Узнав все, о чем мы хотели «допросить» изучаемую клетку, мы перемещаем электрод на долю миллиметра вперед к следующей клетке и снова начинаем всю процедуру тестирования. В любой структуре зрительной системы мы обычно регистрируем реакции сотен клеток в экспериментах, длящихся часы или дни. Раньше или позже у нас начинает складываться некоторое представление о том, что общего имеют клетки данной структуры между собой и в чем они различаются. Поскольку каждая такая структура содержит миллионы клеток, мы можем исследовать лишь малую долю всей нейронной популяции, но, к счастью, различных видов клеток не миллионы, так что рано или поздно мы перестаем находить новые разновидности. Удовлетворившись, мы делаем глубокий вдох и отправляемся на следующий уровень — переходим, например, от наружного коленчатого тела к стриарной коре — и там повторяем всю процедуру. Поведение клеток следующего уровня обычно бывает более сложным, чем на предыдущем уровне, но это различие может быть и совсем небольшим, и весьма существенным. Сравнивая последовательные уровни, мы начинаем понимать вклад каждого из них в анализ нашего зрительного мира — узнаём, какую операцию выполняет каждая из структур над получаемыми ею сигналами, чтобы извлечь из внешней среды биологически полезную для животного информацию. Стриарная кора уже основательно изучена во многих лабораториях. Значительно меньше сведений у нас о следующей корковой зоне — зрительной зоне 2, хотя и здесь мы начинаем получать некоторое представление о том, что делают ее клетки. То же можно сказать и о третьей зоне, средне-височной (MT), которая связана как со стриарной корой, так и со зрительной зоной 2. Дальше, однако, наши познания быстро становятся все более отрывочными: для двух или трех областей мы имеем лишь смутное представление о виде обрабатываемой информации — это может быть, например, анализ цвета или распознавание таких сложных объектов, как человеческие лица; и наконец, примерно о дюжине зон, о которых с уверенностью можно сказать, что они в основном зрительные, мы не знаем практически ничего. Стратегия наша, однако, вполне оправдывается, если судить по быстроте прогресса в этой области. В последующих главах я добавлю некоторые подробности об уровнях вплоть до стриарной коры включительно. В главе 2 я схематично опишу, как «работают» импульсы и синапсы, и приведу ряд примеров нервных путей, иллюстрирующих некоторые общие принципы нейронной организации. Затем мы сосредоточимся на зрении — сначала на анатомии и физиологии сетчатки, а потом на физиологии стриарной коры и ее строении. Далее будут описаны удивительные пространственные отношения нейронов в коре — результат того, что клетки со сходными функциями имеют тенденцию собираться вместе. Затем последует ряд специальных тем: механизмы восприятия цвета и глубины, функция пучка волокон, связывающих два полушария (мозолистого тела), и наконец влияние раннего жизненного опыта на зрительную систему. Некоторые разделы, например о нервном импульсе и цветовом зрении, по необходимости будут содержать несколько больше технических деталей, чем другие. В этих случаях я могу лишь надеяться, что вы последуете мудрому совету: «Если трудно, читай дальше!»
2. Импульсы, синапсы и нейронные сети
Значительную часть нейробиологии составляют разделы о том, как работают отдельные нейроны и как информация передается от клетки к клетке через синапсы. Должно быть очевидным, что без этих сведений мы окажемся в положении человека, желающего понять работу радиоприемника или телевизора, но ничего не знающего о резисторах, конденсаторах и транзисторах. За последние десятилетия благодаря изобретательности ряда нейрофизиологов, из которых наиболее известны Эндрю Хаксли, Алан Ходжкин, Бернард Катц, Джон Экклз и Стивен Куффлер, были хорошо изучены физико-химические механизмы проведения нервных импульсов и синаптической передачи. Однако столь же очевидно, что сведения такого рода сами по себе еще не могут привести к пониманию работы мозга, подобно тому как одни лишь сведения о резисторах, конденсаторах и транзисторах не позволят понять работу радиоприемника или телевизора, а знание химии чернил — прочитать пьесу Шекспира. Эту главу я начинаю с суммирования части того, что нам известно о нервном проведении и синаптической передаче. Большой подмогой в верном понимании существа дела будет знание основ физической химии и электричества, но я думаю, что и без этого читатель сможет получить о предмете достаточное представление. В любом случае для того, чтобы следить за изложением в последующих главах, вам понадобится лишь элементарное понимание этих вопросов. Задача нервной клетки состоит в том, чтобы принимать информацию от клеток, которые ее передают, суммировать, или интегрировать, эту информацию и доставлять интегрированную информацию другим клеткам. Информация обычно передается в форме кратковременных процессов, называемых нервными импульсами. Во всякой клетке каждый импульс бывает точно таким же, как и любой другой, т.е. импульс — это стереотипный процесс. В любой момент частота импульсов, посылаемых нейроном, определяется сигналами, только что полученными им от передающих клеток, и передает информацию клеткам, по отношению к которым этот нейрон является передающим. Частота импульсов варьирует от одного в каждые несколько секунд или еще ниже до максимума около тысячи в секунду.Мембранный потенциал
Что происходит, когда информация передается от одной клетки к другой через синапс? В первой — пресинаптической — клетке около основания аксона возникает электрический сигнал, или импульс. Импульс перемещается по аксону к его окончаниям. Из каждого окончания в результате этого импульса в узкий (0,02 мкм) заполненный жидкостью промежуток, отделяющий одну клетку от другой, — синаптическую щель — высвобождается химическое вещество, которое диффундирует ко второй — постсинаптической — клетке. Оно влияет на мембрану этой второй клетки таким образом, что вероятность возникновения в ней импульсов либо уменьшается, либо возрастает. После этого краткого описания вернемся назад и рассмотрим весь процесс подробно.
 Рис. 8. Схема расположения нервных клеток на поперечном срезе сетчатки, нарисованная Сантьяго Рамон-и-Кахалом, величайшим нейроанатомом всех времен. От верхнего слоя, где показаны более тонкие палочки и более толстые колбочки, до нижнего, где направо выходят волокна зрительного нерва, толщина сетчатки составляет четверть миллиметра.
Рис. 8. Схема расположения нервных клеток на поперечном срезе сетчатки, нарисованная Сантьяго Рамон-и-Кахалом, величайшим нейроанатомом всех времен. От верхнего слоя, где показаны более тонкие палочки и более толстые колбочки, до нижнего, где направо выходят волокна зрительного нерва, толщина сетчатки составляет четверть миллиметра.
Нервная клетка омывается солевым раствором и содержит его внутри. В число солей входит не только хлористый натрий, но также хлористый калий, хлористый кальций и ряд других, менее обычных солей. Поскольку большинство молекул соли диссоциировано, жидкости как внутри, так и снаружи клетки содержат ионы хлора, калия, натрия и кальция (Cl–, K+, Na+ и Ca2+). В состоянии покоя электрические потенциалы внутри и снаружи клетки различаются примерно на одну десятую долю вольта, причем плюс находится снаружи. Точное значение ближе к величине 0,07 вольта, или 70 милливольт. Передаваемые нервами сигналы представляют собой быстрые изменения потенциала, перемещающиеся по волокну от тела клетки к окончаниям аксона. Я начну с описания того, как на клеточной мембране возникает разность потенциалов. Мембрана нервной клетки, покрывающая весь нейрон, — структура чрезвычайно сложная. Она не сплошная, как надувной шарик или шланг, а содержит миллионы «пор», через которые вещества могут переходить с одной стороны на другую. Некоторые из них — это действительно поры различной величины; как сейчас выяснилось, они представляют собой белки в форме трубок, насквозь пронизывающих жировое вещество мембраны. В других случаях это не просто поры, а миниатюрные белковые механизмы, называемые насосами; они способны улавливать ионы одного типа и выбрасывать их из клетки, одновременно захватывая другие ионы внутрь из наружного пространства. Такая перекачка требует затраты энергии, которую клетка в конечном счете получает в процессе окисления глюкозы. Существуют также поры, называемые каналами, — это «клапаны», которые могут открываться и закрываться. Какие воздействия приводят к их открытию или закрытию, зависит от типа пор. На некоторые из них влияет мембранный потенциал, другие открываются или закрываются при наличии определенных веществ во внутренней и наружной жидкости.
 Рис. 9. На этой электронной микрофотографии (срез коры мозжечка крысы) синапс выглядит как узкая темная полоска в нижней части рисунка посередине. Слева от синапса можно видеть поперечное сечение аксона, заполненного мельчайшими круглыми синаптическими пузырьками, в которых хранится нейромедиатор. Справа от синапса виден выступ дендрита (называемый шипиком); он отходит от крупной дендритной ветви, расположенной горизонтально в верхней части рисунка (два темных колбасовидных образования в этом дендрите — митохондрии). В синапсе сближены две мембраны — мембраны аксона и дендрита; здесь они утолщены и выглядят более плотными. Их разделяет щель шириной 20 нанометров.
Рис. 9. На этой электронной микрофотографии (срез коры мозжечка крысы) синапс выглядит как узкая темная полоска в нижней части рисунка посередине. Слева от синапса можно видеть поперечное сечение аксона, заполненного мельчайшими круглыми синаптическими пузырьками, в которых хранится нейромедиатор. Справа от синапса виден выступ дендрита (называемый шипиком); он отходит от крупной дендритной ветви, расположенной горизонтально в верхней части рисунка (два темных колбасовидных образования в этом дендрите — митохондрии). В синапсе сближены две мембраны — мембраны аксона и дендрита; здесь они утолщены и выглядят более плотными. Их разделяет щель шириной 20 нанометров.
Разность потенциалов на мембране в любой момент определяется концентрацией ионов внутри и снаружи, а также тем, открыты или закрыты различные поры. (Выше я говорил, что потенциал влияет на поры, а теперьутверждаю, что поры влияют на потенциал. Давайте пока просто скажем, что эти две вещи могут быть взаимозависимы. Несколько позже будет дано более детальное объяснение.) Так как имеется несколько видов пор и несколько видов ионов, легко понять, что вся система довольно сложна. Когда Ходжкин и Хаксли в 1952 году сумели разобраться в ней, это было огромным достижением. Зададимся прежде всего вопросом: как создается разность потенциалов? Предположим, что вначале никакой разности нет и концентрации ионов внутри и снаружи одинаковы. Пусть далее включается насос, который выводит из клетки ионы одного вида, например натрия, и вместо каждого выведенного иона переносит внутрь ион другого вида, например калия. Сам по себе насос не создает какого-либо потенциала, так как сколько положительно заряженных ионов накачивается внутрь, столько же и откачивается (ионы натрия и калия несут одинаковые положительные заряды). Но представим теперь, что по какой-то причине открылось большое число пор одного типа, например калиевых. Ионы калия начнут перетекать по ним, причем скорость потока через каждую открытую пору будет зависеть от концентрации калия: чем больше ионов возле отверстия поры, тем больше будет их утечка через мембрану; а так как внутри ионов калия больше, чем снаружи, то выходить их будет больше, чем входить внутрь. Но если выходит больше зарядов, чем входит, наружное пространство быстро станет электроположительным по отношению к внутреннему. Это накопление положительного заряда снаружи вскоре начнет противодействовать дальнейшему выходу ионов калия из клетки, так как одноименные заряды отталкивают друг друга. Очень быстро — прежде чем выход ионов K+ приведет к заметному изменению их концентрации — положительный заряд снаружи достигнет величины, при которой он в точности скомпенсирует тенденцию ионов K+ выходить из клетки (с внутренней стороны поры ионов калия больше, но они отталкиваются наружным зарядом). Начиная с этого момента перемещение заряда прекращается, и мы говорим, что система приходит в равновесие. Таким образом, открытие калиевых пор приводит к возникновению на мембране разности потенциалов с положительным полюсом снаружи. Но предположим, что вместо этого открылись натриевые поры. Повторяя все рассуждения с заменой слов «внутренний» на «наружный», вы можете легко убедиться, что результат будет прямо противоположным: снаружи возникнет отрицательный заряд. При одновременном открытии пор того и другого типа результат был бы «компромиссным». Для оценки величины мембранного потенциала мы должны знать относительные концентрации двух ионов и отношение числа открытых и закрытых пор для каждого иона, а затем произвести соответствующие расчеты.
Импульс
Когда нерв находится в покое, большинство калиевых каналов открыто, а большинство натриевых закрыто; поэтому снаружи будет положительный заряд. Во время импульса на коротком отрезке нервного волокна внезапно открывается большое число натриевых каналов, что приводит к кратковременному преобладанию потока ионов натрия, и этот участок быстро становится электроотрицательным снаружи по отношению к внутреннему пространству. Затем натриевые поры вновь закрываются, в то время как калиевые остаются открытыми, причем даже в большем количестве, чем в состоянии покоя. Оба процесса — закрытие натриевых пор и дополнительное открытие калиевых пор — приводят к быстрому восстановлению потенциала покоя с положительным полюсом снаружи. Вся последовательность событий занимает примерно тысячную долю секунды.
 Рис. 10. Вверху: участок аксона в состоянии покоя. Натриевый насос перекачал наружу излишние ионы натрия, а внутрь — недостающие ионы калия. Натриевые каналы в основном закрыты. Поскольку открыто много калиевых каналов, клетку покинуло достаточное количество ионов калия, чтобы мембранный потенциал достиг равновесного в таких условиях уровня — около 70 милливольт с плюсом снаружи.
Внизу: слева направо перемещается нервный импульс. На крайнем правом конце аксон еще пребывает в состоянии покоя. В среднем участке развертываются события, связанные с импульсом: натриевые каналы открыты, ионы натрия переходят внутрь (хотя и не в таком количестве, чтобы их концентрация после одного импульса заметно изменилась); мембранный потенциал 40 милливольт с плюсом внутри. На крайнем левом конце мембрана возвращается в исходное состояние, так как открылись (а затем закрылись) добавочные калиевые каналы, а натриевые каналы автоматически закрылись. Поскольку натриевые каналы не способны сразу же повторно открыться, второй импульс не может возникнуть раньше чем спустя примерно миллисекунду. Это позволяет понять, почему импульс не может повернуть назад к телу клетки.
Рис. 10. Вверху: участок аксона в состоянии покоя. Натриевый насос перекачал наружу излишние ионы натрия, а внутрь — недостающие ионы калия. Натриевые каналы в основном закрыты. Поскольку открыто много калиевых каналов, клетку покинуло достаточное количество ионов калия, чтобы мембранный потенциал достиг равновесного в таких условиях уровня — около 70 милливольт с плюсом снаружи.
Внизу: слева направо перемещается нервный импульс. На крайнем правом конце аксон еще пребывает в состоянии покоя. В среднем участке развертываются события, связанные с импульсом: натриевые каналы открыты, ионы натрия переходят внутрь (хотя и не в таком количестве, чтобы их концентрация после одного импульса заметно изменилась); мембранный потенциал 40 милливольт с плюсом внутри. На крайнем левом конце мембрана возвращается в исходное состояние, так как открылись (а затем закрылись) добавочные калиевые каналы, а натриевые каналы автоматически закрылись. Поскольку натриевые каналы не способны сразу же повторно открыться, второй импульс не может возникнуть раньше чем спустя примерно миллисекунду. Это позволяет понять, почему импульс не может повернуть назад к телу клетки.
Все происходящее зависит от обстоятельств, влияющих на открытие и закрытие пор. Как натриевые, так и калиевые каналы чувствительны к мембранному потенциалу. Уменьшение наружного положительного заряда — деполяризация мембраны относительно состояния покоя — приводит к открытию пор. Это влияние неодинаково для пор двух типов: натриевые поры, открывшись, снова закрываются сами по себе, даже если мембрана остается деполяризованной, и неспособны вновь открыться на протяжении нескольких тысячных долей секунды; калиевые поры остаются открытыми, пока поддерживается деполяризация. При определенном уровне деполяризации число ионов натрия, входящих внутрь, вначале превышает число выходящих ионов калия и наружная поверхность мембраны становится электроотрицательной по отношению к внутренней; позднее начинает преобладать поток калия и восстанавливается потенциал покоя. В этой последовательности событий, составляющих импульс (открываются поры, ионы проходят через мембрану и мембранный потенциал дважды претерпевает изменения), число ионов, фактически проходящих через мембрану — Na+ внутрь, а K+ наружу, — ничтожно, и его недостаточно для измеримого изменения ионных концентраций внутри или снаружи клетки. За несколько минут, однако, нейрон способен разрядиться тысячу раз, и в результате концентрации ионов могли бы заметно измениться, если бы не насос, который все время выводит натрий и накачивает внутрь калий, поддерживая таким образом их концентрации на надлежащих уровнях покоя. Почему во время импульса столь малый перенос заряда ведет к таким большим колебаниям потенциала? Это следствие одного из простых законов электричества: емкость мембраны мала, а потенциал равен перенесенному заряду, деленному на емкость. Деполяризация мембраны — уменьшение электроотрицательности внутри относительно состояния покоя — вот что обеспечивает первоначальный запуск импульса. Если быстро ввести в покоящееся волокно некоторое количество ионов натрия, вызвав небольшую начальную деполяризацию, то в результате откроется небольшое число натриевых пор; но, поскольку много калиевых пор уже открыто, изнутри может выйти достаточно калия, чтобы скомпенсировать этот эффект и быстро вернуть мембрану в исходное состояние покоя. Предположим, однако, что начальный перенос заряда столь велик и открылось так много натриевых пор, что натрий приносит внутрь больше заряда, чем может быть выведено с калием; тогда мембрана деполяризуется еще сильнее. Это приведет к открытию еще большего числа натриевых пор, к еще большей деполяризации и так далее — возникнет самоусиливающийся взрывоподобный процесс. Когда откроются все натриевые поры, которые могут открыться, мембранный потенциал изменит свой знак на обратный по отношению к потенциалу покоя: вместо 70 милливольт с положительным полюсом снаружи он составит 40 милливольт с отрицательным полюсом снаружи. Уменьшение потенциала на мембране с последующим изменением его знака (реверсией) не происходит сразу по всей длине волокна, так как перенос заряда требует времени. Активный участок возникает в одном месте и перемещается по волокну со скоростью от 0,1 до примерно 10 метров в секунду. В любой момент времени существует один активный участок с реверсированным потенциалом, и эта область реверсии передвигается, удаляясь от тела нейрона; впереди нее находится участок с еще не открытыми каналами, а позади — участок, где каналы снова закрылись и временно неспособны к повторному открытию. Это и есть процесс распространения импульса. Вы видите, что он вовсе не похож на прохождение тока по медной проволоке. По всей длине нерва не перемещаются электрические заряды, ионы или вообще нечто материальное, точно так же как при смыкании лезвий ножниц ничто не перемещается от соединительного винта до их кончиков. (Ионы образуют лишь местные токи, переходя внутрь и наружу, подобно тому как лезвия ножниц движутся вверх и вниз.) Перемещается некоторое событие или процесс — перекрещивание лезвий ножниц или импульс в нерве. Поскольку подготовка натриевых каналов к следующему открытию и закрытию требует некоторого времени, наибольшая частота, с которой клетка или аксон способны генерировать импульсы, составляет около 800 в секунду. Однако столь высокая частота необычна; даже для сильно активированных нервных волокон характерны частоты 100–200 импульсов в секунду.
 Рис. 11. Мембрана глиальной клетки многократно обернута вокруг аксона, как это видно на электронной микрофотографии поперечного среза нервного волокна. Такая мембрана состоит из миелина, который ускоряет проведение нервных импульсов, повышая сопротивление и уменьшая емкость между внутренностью аксона и окружающим пространством. В аксоне видны (в поперечном сечении) органеллы, называемые микротрубочками.
Рис. 11. Мембрана глиальной клетки многократно обернута вокруг аксона, как это видно на электронной микрофотографии поперечного среза нервного волокна. Такая мембрана состоит из миелина, который ускоряет проведение нервных импульсов, повышая сопротивление и уменьшая емкость между внутренностью аксона и окружающим пространством. В аксоне видны (в поперечном сечении) органеллы, называемые микротрубочками.
Важной особенностью нервного импульса является то, что он возникает по принципу всё или ничего. При достаточной начальной деполяризации — если она превышает некоторую пороговую величину — процесс становится самоусиливающимся и реверсия происходит всегда до 0,02 вольт (минус снаружи). Величина потенциала, распространяющегося по нерву (т.е. импульса), определяется самим нервом, а не степенью начальной деполяризации, которая привела к его возникновению. Здесь уместна аналогия с любым взрывообразным процессом. Скорость полета пули никак не связана с тем, насколько резко вы нажали на спусковой крючок. Для многих функций мозга скорость проведения импульса представляется весьма важной, и нервная система выработала особый механизм ее повышения. Плазматическая мембрана глиальных клеток многократно обертывается вокруг аксона, образуя слоистую оболочку, значительно повышающую эффективную толщину нервной мембраны. Это утолщение снижает емкость мембраны, а тем самым и величину заряда, необходимую для ее деполяризации. Слоистое вещество, богатое жировым материалом, называется миелином. Через каждые несколько миллиметров оболочка прерывается в так называемых перехватах Ранвье, что позволяет токам, связанным с импульсом, входить в аксон или выходить из него. В результате нервный импульс фактически перескакивает от одного перехвата к следующему, а не передвигается непрерывно вдоль аксона, что намного ускоряет передачу нервных сигналов. Большинство крупных волокнистых пучков в мозгу миелинизировано, что придает им блестящий белый цвет на свежеприготовленных срезах. Белое вещество головного и спинного мозга состоит из миелинизированных аксонов, и в нем отсутствуют тела нервных клеток, дендриты и синапсы. Серое вещество состоит в основном из клеточных тел, дендритов, аксонных окончаний и синапсов, но может содержать и миелинизированные аксоны. Основные пробелы в нашем понимании природы импульса, равно как и основные направления современных исследований в этой области, связаны со структурой и функцией белковых каналов.
Синаптическая передача
Как первоначально возникают импульсы, и что происходит на дальнем конце аксона, когда импульс прибывает туда? Участок клеточной мембраны у окончания аксона, образующий первую половинку синапса (пресинаптическую мембрану), обладает удивительной специализированной структурой. Прежде всего, он содержит особые каналы, которые при деполяризации открываются и пропускают положительно заряженные ионы кальция. Поскольку концентрация кальция (как и натрия) снаружи выше, чем внутри клетки, открытие этих каналов позволяет кальцию переходить внутрь. Каким-то пока не известным образом это поступление кальция внутрь клетки приводит к выбрасыванию через мембрану наружу небольших порций особых веществ, называемых нейромедиаторами. Уже идентифицировано около двадцати химических медиаторов, и, судя по темпу новых открытий, их общее число может превышать полсотни. Молекулы медиаторов намного меньше белковых молекул, но обычно крупнее ионов натрия или кальция. Примерами нейромедиаторов могут служить ацетилхолин и норадреналин. Когда эти вещества высвобождаются из пресинаптической мембраны, они быстро диффундируют через синаптическую щель шириной 0,02 мкм к постсинаптической мембране. Постсинаптическая мембрана тоже специализирована: в ней имеются белковые рецепторы, которые реагируют на нейромедиатор открытием соответствующих каналов, позволяя ионам одного или нескольких типов проходить через них. От того, какие именно ионы (натрий, калий, хлор) смогут проходить, зависит, будет ли сама постсинаптическая клетка деполяризована или же ее мембранный потенциал будет стабилизирован, т.е. ее деполяризация затруднена. Подытожим сказанное. Нервный импульс приходит к окончанию аксона и вызывает здесь высвобождение специальных нейромедиаторных молекул. Эти нейромедиаторы воздействуют на постсинаптическую мембрану так, что либо понижают ее мембранный потенциал, либо предотвращают его понижение. При понижении мембранного потенциала частота импульсации возрастает; мы назовем такой синапс возбуждающим. Если же мембранный потенциал вместо этого стабилизируется на подпороговом уровне, импульсы не возникают или возникают с меньшей частотой, и тогда синапс называют то́рмозным. Будет ли данный синапс возбуждающим или тормозным, зависит от того, какой в нем высвобождается медиатор и каковы здесь рецепторные молекулы. Ацетилхолин, самый известный медиатор, в некоторых синапсах оказывает возбуждающее действие, а в других — тормозное, он возбуждает мышцы конечностей и туловища, но тормозит сокращения сердца. Норадреналин обычно служит возбуждающим медиатором, гамма-аминомасляная кислота (ГАМК) — тормозящим. Насколько нам известно, характер действия каждого данного синапса остается постоянным на протяжении всей жизни животного. С дендритами и телом нейрона могут контактировать десятки, сотни или тысячи аксонных окончаний; поэтому в любой момент одни входные синапсы стремятся деполяризовать клетку, а другие противодействуют этому. Импульс, приходящий к возбуждающему синапсу, будет деполяризовать постсинаптическую клетку; если одновременно придет также импульс к тормозному синапсу, эффекты обоих импульсов будут стремиться погасить друг друга. В любой момент уровень мембранного потенциала является результатом сложения вместе возбуждающих и тормозных влияний. Одиночный импульс, приходящий к одному синапсу, обычно лишь очень слабо воздействует на постсинаптическую клетку, и его эффект через несколько миллисекунд исчезает. При поступлении импульсов от нескольких других нейронов клетка суммирует, или интегрирует, их влияния. При достаточном понижении мембранного потенциала — если возбуждающие сигналы приходят к достаточному числу синапсов и с достаточно высокой частотой — суммарная деполяризация сможет привести к возникновению импульсов, обычно в виде целой их серии. Импульсы обычно возникают в том месте, где от тела клетки отходит аксон: деполяризация данной величины здесь с наибольшей вероятностью может вызвать импульс — по-видимому, благодаря особенно высокой плотности расположения натриевых каналов в мембране. Чем сильнее в этом месте деполяризована мембрана, тем больше импульсов возникает каждую секунду. Почти все клетки нервной системы имеют входные синапсы от нескольких других клеток. Это называют конвергенцией. В то же время аксоны почти всех клеток многократно ветвятся и обслуживают большое число других нейронов — возможно, сотни или тысячи. Мы называем это дивергенцией. Легко видеть, что без конвергенции и дивергенции нервная система немногого бы стоила: возбуждающий синапс просто передавал бы каждый импульс следующей клетке, не выполняя никакой полезной функции, а тормозному синапсу, который был бы единственным входом данной клетки, нечего было бы тормозить, если только постсинаптическая клетка не обладала бы каким-то специальным механизмом, заставляющим ее разряжаться спонтанно. Мне хотелось бы сделать заключительное замечание о сигналах, передаваемых нервными волокнами. Хотя аксоны почти всегда проводят импульсы по принципу «всё или ничего», имеются некоторые исключения. Если локальная деполяризация оказывается подпороговой, т.е. недостаточной для запуска взрывоподобного импульса типа «все или ничего», она тем не менее имеет некоторую тенденцию распространяться вдоль волокна, уменьшаясь со временем и с расстоянием от начального пункта. (При обычном проведении нервного импульса именно такое локальное распространение и доводит потенциал соседнего, покоящегося участка нервной мембраны до пороговой деполяризации, при которой начинается самоусиливающийся процесс.) Некоторые аксоны столь коротки, что не возникает нужды в распространяющихся импульсах: деполяризация клеточного тела или дендритов путем пассивного распространения способна создавать в окончании аксона деполяризацию, достаточную для высвобождения медиатора. У млекопитающих известные случаи передачи информации без импульсов немногочисленны, но важны. В наших сетчатках нервные клетки двух или трех из пяти типов функционируют без импульсов. Важное отличие этих пассивно передаваемых сигналов от импульсов (помимо их малой и прогрессивно убывающей амплитуды) состоит в том, что их величина меняется в зависимости от силы стимула. Поэтому их часто называют градуальными сигналами. Чем больше сигнал, тем сильнее деполяризация окончания и тем больше высвобождается медиатора. Следует напомнить, что импульсы, напротив, не увеличиваются по амплитуде с ростом стимула — вместо этого возрастает частота их повторения. И чем чаще возникают импульсы, тем больше медиатора выделяется в окончаниях. Таким образом, конечный результат не слишком различен. Нередко говорят, что градуальные потенциалы представляют собой пример аналоговых сигналов, а импульсы, будучи событиями типа «все или ничего», носят характер цифровой сигнализации. Я нахожу эту аналогию неверной, поскольку точное положение каждого импульса в серии в большинстве случаев не имеет никакого значения. Что важно, так это среднее число их в данном временном интервале, а не тонкие детали. Таким образом, оба вида сигналов существенно аналоговые.
Типичный нейронный путь
Теперь, когда мы кое-что знаем об импульсах, синапсах, возбуждении и торможении, можно задаться вопросом, как из нейронов строятся более крупные образования. Мы можем представить центральную нервную систему — головной и спинной мозг — как некоторый ящик со входом и выходом. Входные сигналы оказывают воздействие на особые нервные клетки, называемые рецепторами; эти клетки реагируют не на синаптические входы от других клеток, а на то, что мы можем обозначить нестрогим термином «внешняя информация». Эта информация может принимать форму света, проникающего в наши глаза; механической деформации нашей кожи, барабанной перепонки или полукружных каналов; химических веществ, как в наших органах обоняния или вкуса. Во всех этих случаях под влиянием стимула в рецепторах возникает электрический сигнал и в результате изменяется скорость высвобождения медиатора в окончаниях их аксонов. (Вас не должно смущать двойное значение термина рецептор; вначале он означал клетку со специализированной реакцией на сенсорные стимулы, но впоследствии был применен также к белковым молекулам со специализированной реакцией на нейромедиаторы.) На другом конце нервной системы мы имеем выход — двигательные нейроны, отличающиеся от всех остальных тем, что их аксоны оканчиваются не на других нейронах, а на мышечных клетках. Все выходные сигналы нашей нервной системы вызывают мышечные сокращения, за редким исключением сигналов, адресуемых клеткам желез. Это, по существу, единственный способ, которым мы можем оказывать влияние на окружающую нас среду. Отключите у какого-либо животного мышцы, и вы полностью отрежете его от остального мира; отключите вместо этого входные сигналы — и прекратятся все внешние влияния, что опять-таки превратит животное фактически в растение. Согласно одному из возможных определений, животное — это организм, реагирующий на внешние события и своими действиями оказывающий влияние на внешний мир.
 Рис. 12. Нервно-мышечное соединение у лягушки. Более тонкое нервное волокно извивается рядом с двумя мышечными волокнами, образуя синапс в нижней левой части снимка.
Рис. 12. Нервно-мышечное соединение у лягушки. Более тонкое нервное волокно извивается рядом с двумя мышечными волокнами, образуя синапс в нижней левой части снимка.
Центральная нервная система, лежащая между входными и выходными нейронами, является тем аппаратом, который позволяет нам воспринимать, реагировать и помнить; он же в конечном счете должен быть ответствен за наше сознание, совесть и душу. Одна из главных целей нейробиологии состоит в выяснении того, что происходит на этом пути — как информация, приходящая к некоторой группе нейронов, преобразуется и затем передается дальше, а также какой смысл имеют эти преобразования для успешной адаптации животного к среде. Хотя схемы связей для многих отделов центральной нервной системы значительно варьируют в деталях, в большинстве случаев в основе их лежит сравнительно простой общий план, показанный на рис. 13. Приведенная здесь схема — это скорее карикатура, которую не следует понимать буквально и которая требует уточнений, обсуждаемых ниже. Слева на рисунке показаны рецепторы — ряд преобразующих информацию нейронов, каждый из которых обслуживает один из видов ощущений, например осязание, вибрационное чувство или зрительное восприятие. Мы можем считать эти рецепторы первым уровнем какого-то сенсорного пути. Волокна от рецепторов образуют синаптические контакты со вторым рядом нервных клеток — вторым уровнем нашей схемы; эти клетки в свою очередь образуют контакты с третьим уровнем и так далее. «Уровень» не является техническим или широко применяемым нейроанатомическим термином, но мы еще увидим его полезность. Иногда три или четыре таких уровня объединяются вместе в более крупный элемент, который я за недостатком лучшего или общепринятого термина назову структурой. Структуры — это скопления клеток, обычно в виде пластин или сферических ядер, упоминавшихся в главе 1. В случае пластинчатой структуры каждый из образующих ее уровней может быть дискретным слоем клеток. Хорошим примером служит сетчатка, в которой три таких слоя и, грубо говоря, три уровня. Если несколько уровней группируются, образуя более крупную структуру, то нервные волокна, приходящие к ней от предыдущей структуры и идущие от нее к следующей, обычно группируются в пучки, называемые трактами. На схеме (рис. 13) можно видеть, насколько распространены процессы дивергенции и конвергенции сигналов: аксон почти каждой клетки данного уровня ветвится при подходе к следующему уровню и оканчивается на нескольких или многих клетках; и наоборот, клетка любого уровня, кроме первого, имеет синаптические входы от нескольких или многих клеток предыдущего уровня.
 Рис. 13. Многие отделы центральной нервной системы организованы в виде последовательных слоев-уровней. Клетка одного уровня получает многочисленные возбуждающие и тормозные входы от предыдущего уровня и посылает выходные сигналы многим клеткам последующего уровня. Основную массу входной информации нервная система получает от рецепторов глаз, ушей, кожи и т.д., которые преобразуют такие внешние воздействия, как свет, тепло или звук, в электрические нервные сигналы. Выходом могут быть сокращения мышц или реакции железистых клеток.
Рис. 13. Многие отделы центральной нервной системы организованы в виде последовательных слоев-уровней. Клетка одного уровня получает многочисленные возбуждающие и тормозные входы от предыдущего уровня и посылает выходные сигналы многим клеткам последующего уровня. Основную массу входной информации нервная система получает от рецепторов глаз, ушей, кожи и т.д., которые преобразуют такие внешние воздействия, как свет, тепло или звук, в электрические нервные сигналы. Выходом могут быть сокращения мышц или реакции железистых клеток.
Очевидно, что мы должны усовершенствовать и конкретизировать эту упрощенную схему, и у нас для этого есть некоторая отправная модель. Прежде всего на входном конце мы имеем не одну, а много сенсорных систем — зрение, осязание, вкус, обоняние и слух, и каждая система имеет свой собственный набор уровней в мозгу. Когда и где в мозгу различные наборы уровней сводятся вместе, если они действительно сводятся, все еще неясно. Прослеживая одну систему, например зрительную или слуховую, от рецепторов до мозга, мы можем обнаружить, что она разделяется на отдельные подсистемы. В случае зрения эти подсистемы имеют отношение специально к движениям глаз, сокращению зрачка, восприятию формы, движения, глубины или цвета объектов. Таким образом, вся система дивергирует на отдельные каналы (пути); таких каналов может быть много и они могут сильно разниться по своей длине. Уже сразу видно, что некоторые пути содержат много структур на своем протяжении, а другие — мало. При более детальном анализе оказывается, что какой-то аксон одного уровня может идти дальше, не заходя на один или два последующих уровня сенсорной системы, и даже прямо направиться к двигательному нейрону. (Такой пропуск уровней в нейроанатомии аналогичен тому, что часто случается в генеалогии. Современный английский монарх не связан с Вильгельмом Завоевателем однозначным числом поколений: число приставок «пра» к слову «дед» неопределенно из-за родственных браков между племянниками и тетками и других еще более сомнительных обстоятельств.) Если путь от входа к выходу очень короток, мы называем это рефлексом. В зрительной системе сужение зрачка при ярком свете служит примером рефлекса, в котором число синапсов, вероятно, около шести. В наиболее простом случае аксон от рецептора оканчивается прямо на двигательном нейроне, так что мы имеем от входа к выходу всего три клетки (рецептор, двигательный нейрон и мышечное волокно) и только два синапса; такую ситуацию мы называем моносинаптической рефлекторной дугой. (Возможно, человек, сочинивший этот термин, не рассматривал нервно-мышечное соединение как настоящий синапс или не умел считать до двух.) Этот короткий путь активируется, когда врач стучит молоточком по вашему колену и нога подпрыгивает. Джон Николлс обычно говорил своей аудитории в Гарвардской медицинской школе, что есть только два резона проверять этот рефлекс: убить время и проверить, нет ли у больного сифилиса. На выходном конце мы находим не только различные группы мышц, расположенные на туловище, в конечностях, глазах и языке, которыми мы можем управлять произвольно, но также мускулатуру, обслуживающую менее произвольные или непроизвольные, «хозяйственные» функции, такие как взбалтывание содержимого желудка, мочеиспускание или дефекация, а в промежутках между этими актами — закрытие выводных протоков с помощью сфинктеров. Мы должны также дополнить нашу модель путями обратной передачи информации. На нашей схеме (рис. 13), разумеется, доминирует направление слева направо, от входа к выходу. Но почти во всех случаях, когда сигналы передаются от одного уровня к следующему, возвратные связи направляют их также назад, со второго уровня на первый. (Иногда мы можем догадываться, для чего такая обратная связь могла бы служить, но почти во всех случаях у нас нет на этот счет ясного понимания.) Наконец, даже в пределах какого-то одного уровня мы часто обнаруживаем богатую сеть соединений между соседними клетками. Таким образом, утверждение, что та или иная структура содержит определенное число уровней, почти всегда будет чрезмерным упрощением. Когда я начинал работать в области неврологии в начале 50-х годов, этот основной план нервной системы был хорошо известен. Но в те дни никто не имел четкого представления о том, как понимать организацию этой информационной эстафеты по цепи уровней. Сейчас мы значительно больше знаем о способах преобразования информации в некоторых частях мозга; о других частях нам еще почти ничего не известно. Оставшиеся главы этой книги посвящены зрительной системе, которую мы сегодня понимаем лучше всего. Ниже я попытаюсь дать обзор некоторых фактов, известных нам об этой системе.
Зрительный путь
Теперь мы можем приспособить нашу первоначальную схему (рис. 13) к специальному случаю зрительного пути. Как показано на рис. 14, рецепторы и следующие два уровня находятся в сетчатке. Рецепторами служат палочки и колбочки; зрительный нерв — общий выход всей сетчатки — представляет собой пучок аксонов клеток третьего уровня, называемых ганглиозными клетками сетчатки. Между рецепторами и ганглиозными клетками имеются промежуточные нейроны, из которых наиболее важны биполярные клетки. Зрительный нерв направляется к расположенной глубоко в мозгу промежуточной станции — наружному коленчатому телу. Отсюда после всего лишь одного набора синапсов выходные сигналы направляются к стриарной коре, содержащей три или четыре уровня.
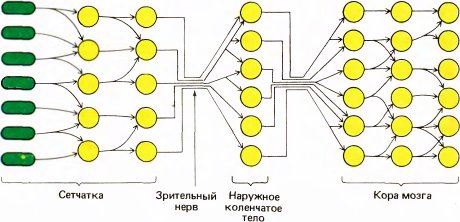 Рис. 14. Начальные уровни зрительной системы млекопитающих имеют слоистую организацию, часто встречающуюся в центральной нервной системе. Первые три уровня размещаются в сетчатке, остальные в мозгу — в наружных коленчатых телах и далее в коре мозга.
Рис. 14. Начальные уровни зрительной системы млекопитающих имеют слоистую организацию, часто встречающуюся в центральной нервной системе. Первые три уровня размещаются в сетчатке, остальные в мозгу — в наружных коленчатых телах и далее в коре мозга.
 Рис. 15. Любой уровень в схемах на рис. 13 и 14 состоит из двумерного слоя клеток. Клетки любого уровня могут быть так плотно упакованы, что им приходится размещаться в несколько слоев; тем не менее они все принадлежат к одному уровню.
Рис. 15. Любой уровень в схемах на рис. 13 и 14 состоит из двумерного слоя клеток. Клетки любого уровня могут быть так плотно упакованы, что им приходится размещаться в несколько слоев; тем не менее они все принадлежат к одному уровню.
Каждую колонку приведенной схемы можно представить себе как пластину из клеток в поперечном разрезе. Если, например, вы будете смотреть налево от слоя, расположенного на странице справа, то увидите все клетки предыдущего слоя. Каждая клеточная колонка на рисунке представляет собой двумерный массив клеток, подобно тому как это показано для палочек и колбочек на рис. 15. Начиная здесь рассуждать об отдельных уровнях, мы сразу же должны вспомнить о нашем сравнении с генеалогией. В сетчатке, как мы увидим в главе 3, минимальное число уровней от рецепторов до выхода составляет, несомненно, три, но из-за наличия двух других типов клеток некоторая часть информации проходит непрямыми путями с четырьмя или пятью уровнями от входа до выхода. Для удобства на схеме эти окольные пути, несмотря на их важность, не показаны; кроме того, упрощены также связи между нейронами. Когда я говорю о ганглиозных клетках сетчатки как об «уровне 3 или 4», это не значит, что я забыл, сколько их есть. Для того чтобы понять характер передачи сигналов в подобного рода сети, мы можем для начала рассмотреть поведение одиночной ганглиозной клетки сетчатки. Из анатомии мы знаем, что такая клетка имеет входы от многих биполярных клеток — возможно от 12, 100 или 1000, каждая из которых тоже в свою очередь получает сигналы от сходного числа рецепторов. Как общее правило, все клетки, посылающие сигналы к какой-то одной клетке данного уровня (как, например, биполярные клетки, «питающие» одну ганглиозную клетку), тесно группируются вместе. В сетчатке нейроны, соединенные с какой-либо одной клеткой следующего уровня, обычно лежат в зоне диаметром от 1 до 2 миллиметров, и уж во всяком случае не разбросаны по всей сетчатке. Иными словами, внутри сетчатки нет соединений длиннее примерно 1–2 миллиметров. Если бы мы располагали подробным описанием всех связей в такой структуре и достаточными познаниями в клеточной физиологии — например, знали, какие синапсы возбуждающие, а какие тормозные, то мы в принципе могли бы вывести заключение о происходящих здесь преобразованиях информации. Однако в отношении сетчатки и коры мозга имеющиеся в нашем распоряжении сведения отнюдь не удовлетворяют этому условию. Поэтому наиболее эффективный подход к решению этой задачи заключался до сих пор в микроэлектродной регистрации активности клеток и в сопоставлении их входных сигналов и выходных реакций. В зрительной системе это равносильно поискам ответа на вопрос: что происходит в клетке, будь то ганглиозная клетка сетчатки или клетка коры, когда глазу предъявляют тот или иной зрительный стимул. При попытке активировать светом клетку уровня 3 (ганглиозную), нашим первым интуитивным поступком, вероятно, было бы осветить все питающие ее палочки и колбочки ярким светом, направленным в глаз. Именно так полагало большинство исследователей конца 40-х годов, когда физиологи еще только начинали осознавать важность синаптического торможения и никто не понимал, что тормозные синапсы столь же многочисленны, как и возбуждающие. Из-за торможения результат всякой стимуляции решающим образом зависит от того, куда в точности попадает свет, и от того, какие связи являются тормозными, а какие возбуждающими. Если мы хотим сильно активировать ганглиозную клетку, то едва ли не самое глупое, что можно сделать, это стимулировать все соединенные с ней палочки и колбочки. Стимуляция большим световым пятном или, как предельный случай, освещение всей сетчатки рассеянным светом обычно приводит к тому, что импульсация клетки не учащается и не замедляется, т.е. фактически ничего не происходит: клетка просто сохраняет импульсацию на уровне покоя порядка 5–10 импульсов в секунду. Для повышения частоты разрядов мы должны освещать некоторое частичное подмножество рецепторов, а именно те из них, которые связаны с данной клеткой (через биполярные нейроны) таким образом, что их влияние окажется возбуждающим. Освещение только одного такого рецептора едва ли может дать сколько-нибудь заметный эффект, но если бы мы осветили все рецепторы с возбуждающим эффектом, то мы были бы вправе ожидать суммирования их влияний и активации клетки — именно так фактически и происходит. Как мы увидим позже, наилучшим стимулом для большинства ганглиозных клеток сетчатки оказывается небольшое световое пятнышко строго определенного размера, вспыхивающее в строго определенном месте. Это, кроме всего прочего, говорит о том, какую важную роль в функции сетчатки играет торможение.
Произвольное движение
Хотя книга будет посвящена в основном начальным, сенсорным уровням нервной системы, я хочу привести два примера, касающихся движения, чтобы дать представление о том, что могут делать конечные уровни схемы на рис. 13. Рассмотрим сначала, как движутся наши глаза. Каждый глаз имеет почти сферическую форму и может свободно поворачиваться подобно шарику в гнезде. (Если бы глазу не надо было двигаться, он вполне мог бы возникнуть в процессе эволюции в виде прямоугольного ящика, похожего на старинный фотографический аппарат.) К каждому глазу прикреплены шесть наружных глазных мышц, которые управляют его движениями. Для нас здесь не существенно, как все эти мышцы прикрепляются к глазу, но из рис. 16 легко видеть, что для поворота одного из глаз, скажем правого, внутрь к носу человек должен расслабить наружную прямую мышцу и сократить внутреннюю прямую. Если бы каждая мышца не обладала все время некоторым тонусом, глаз бы свободно болтался в глазнице; поэтому любое движение глаза осуществляется путем сокращения одной мышцы и в точности такого же по величине расслабления мышцы-антагониста. (Это относится и к работе почти всех мышц тела.) Далее, всякое движение одного глаза почти всегда является частью более сложного комплекса движений. Если мы смотрим на объект, расположенный очень близко, оба глаза поворачиваются внутрь; если мы смотрим налево, правый глаз поворачивается внутрь, а левый — наружу; если же мы смотрим вверх или вниз, оба глаза вместе поворачиваются вверх или вниз.
 Рис. 16. Положение каждого глаза контролируется шестью отдельными мышцами, две из которых показаны на этом рисунке. Эти мышцы — наружная прямая и внутренняя прямая — контролируют горизонтальный поворот глаз, осуществляемый при переводе взгляда слева направо или от близких объектов к удаленным. Другие восемь мышц, по четыре на каждый глаз, контролируют поднятие и опускание глаз, т.е. поворот в вертикальной плоскости.
Рис. 16. Положение каждого глаза контролируется шестью отдельными мышцами, две из которых показаны на этом рисунке. Эти мышцы — наружная прямая и внутренняя прямая — контролируют горизонтальный поворот глаз, осуществляемый при переводе взгляда слева направо или от близких объектов к удаленным. Другие восемь мышц, по четыре на каждый глаз, контролируют поднятие и опускание глаз, т.е. поворот в вертикальной плоскости.
Всеми этими движениями управляет мозг. Каждую глазную мышцу заставляет сокращаться импульсация двигательных (моторных) нейронов, расположенных в участке мозга, который называют мозговым стволом. Каждой из двенадцати мышц соответствует небольшое скопление из нескольких сотен двигательных нейронов ствола. Эти скопления называются глазодвигательными ядрами. Каждый моторный нейрон глазодвигательного ядра обслуживает несколько мышечных волокон одной из глазных мышц. Эти нейроны в свою очередь получают сигналы от других возбуждающих волокон. Для такого движения, как конвергенция глаз, нужно, чтобы эти предшествующие нейроны посылали ответвления своих аксонов к соответствующим двигательным нейронам, обслуживающим обе внутренние прямые мышцы. Такая одиночная предшествующая клетка могла бы иметь ветвящийся надвое аксон, одна ветвь которого идет к одному глазодвигательному ядру, а другая — к его двойнику на противоположной стороне. В то же самое время нужно, чтобы другая предшествующая нервная клетка (или клетки), аксон которой имеет тормозящие окончания на двигательных нейронах наружной прямой мышцы, вызывала ее строго соразмерное расслабление. Необходимо, чтобы оба набора предшествующих клеток разряжались совместно, обеспечивая одновременное сокращение и расслабление, а для этого мы могли бы иметь одну управляющую клетку (или группу клеток) на еще более раннем уровне, которая бы возбуждала обе группы. Таков один из механизмов, позволяющих получить координированные движения с участием многих мышц. Практически каждое осуществляемое нами движение — результат совместного сокращения многих мышц и расслабления многих других. Если вы захотите сжать пальцы в кулак, то мышцы передней поверхности вашего предплечья (на той стороне руки, где ладонь) сократятся, что можно ощутить, положив на предплечье другую руку. (Большинство людей, наверное, думают, что мышцы, сгибающие пальцы, находятся в кисти руки. Кисть действительно содержит некоторые мышцы, но это не сгибатели пальцев.) Как показано на рис. 17, мышцы предплечья, сгибающие пальцы, связаны с тремя косточками каждого пальца длинными сухожилиями, ход которых можно проследить на передней поверхности запястья. Некоторое удивление может вызвать то, что при сжимании кулака сокращаются также мышцы на тыльной стороне предплечья. Это может показаться совершенно излишним, если не учитывать, что, сжимая кулак, мы хотим жестко зафиксировать запястье в среднем положении: если бы сократились только сгибатели пальцев, их сухожилия, проходя по передней поверхности запястья, согнули бы также и его. Нужно скомпенсировать эту тенденцию к нежелательному сгибанию запястья путем сокращения мышц, отгибающих запястье назад, а они находятся как раз на тыльной стороне предплечья. Важно то, что мы это делаем не думая. Более того, мы не обучаемся этому, посещая утренние лекции или оплачивая услуги тренера. Новорожденный ребенок хватает вас за палец и крепко держится за него настоящим кулаком без какой-либо предварительной тренировки. По-видимому, в спинном мозгу у нас есть какие-то клетки исполнительного типа, которые посылают возбуждающие ветви как к сгибателям пальцев, так и к разгибателям запястья, и функция которых состоит в обслуживании процесса сжатия кулака. Видимо, связи этих клеток полностью «запаяны» еще до рождения, так же как и связи клеток, которые позволяют сводить глаза при разглядывании близких предметов, не думая об этом и не обучаясь этому.
 Рис. 17. При сжатии пальцев в кулак действуют мышцы, сухожилия которых проходят в кисть впереди от лучезапястного сустава и поэтому стремятся согнуть также и кисть. Чтобы противодействовать этому и фиксировать кисть, должны сокращаться и разгибатели кисти.
Рис. 17. При сжатии пальцев в кулак действуют мышцы, сухожилия которых проходят в кисть впереди от лучезапястного сустава и поэтому стремятся согнуть также и кисть. Чтобы противодействовать этому и фиксировать кисть, должны сокращаться и разгибатели кисти.
3. Глаз
Глаз часто сравнивают с фотоаппаратом. Более уместно было бы сравнить его с телевизионной камерой, установленной на треноге, с автоматической системой слежения — машиной, которая самофокусируется, автоматически подстраивается к интенсивности света, имеет самоочищающуюся линзу и присоединена к компьютеру со столь развитыми возможностями параллельной обработки информации, что инженеры еще только начинают обсуждать сходные стратегии для конструируемой ими аппаратуры. Гигантская работа по преобразованию света, падающего на две сетчатки, в осмысленную зрительную сцену часто странным образом игнорируется, как будто все необходимое нам для того, чтобы видеть, — это изображение внешнего мира, четко сфокусированное на сетчатке. Хотя получение резких изображений и важная задача, она скромна по сравнению с работой нервной системы — сетчатки и мозга. Как мы увидим в этой главе, вклад сетчатки уже сам по себе впечатляет. Преобразуя свет в нервные сигналы, она начинает извлекать из окружающей среды то, что полезно, и отбрасывать то, что излишне. Никакое человеческое изобретение, включая управляемые компьютером камеры, пока еще не может соперничать с глазом. Эта глава посвящена главным образом нейронной части глаза — сетчатке, но я начну с краткого описания глазного яблока, аппарата, который содержит сетчатку и создает на ней четкое изображение внешнего мира.Глазное яблоко
Совокупная функция несетчаточных частей глаз заключается в том, чтобы обеспечить на двух сетчатках сфокусированное четкое изображение внешнего мира. Каждый глаз устанавливается в глазнице в определенное положение шестью маленькими наружными мышцами, упоминавшимися в главе 2. То, что каждый глаз имеет именно шесть мышц, не случайно; они разбиваются на три пары, причем мышцы каждой пары работают в противофазе, обеспечивая движения в трех ортогональных (перпендикулярных) плоскостях. Для обоих глаз задача слежения за объектом должна выполняться с точностью до нескольких угловыхминут — иначе видимое изображение будет двоиться. (Чтобы понять, насколько такое двоение может быть мучительным, попробуйте посмотреть на что-нибудь, надавив на край одного из глаз указательным пальцем.) Столь точные движения требуют для своей реализации набора тонко настроенных рефлексов, включая те, которые контролируют положение головы. Роговица (прозрачная передняя часть глаза) и хрусталик вместе образуют эквивалент линзы фотоаппарата. Приблизительно две трети общего преломления света, необходимого для фокусировки, происходит на границе воздух — роговица, где свет входит в глаз. Оставшуюся треть фокусирующей способности реализует хрусталик, но его главная задача — обеспечить необходимое регулирование для фокусировки на объектах, расположенных на разных расстояниях от глаза. Фокусируя фотоаппарат, мы изменяем расстояние от линзы до фотопленки; в глазу же изменяется не расстояние от хрусталика до сетчатки, а форма эластичного студенистого хрусталика — путем натяжения или ослабления прикрепленных к его краю сухожилий таким образом, что для близких объектов он делается более выпуклым, а для удаленных — более плоским. Эти изменения формы осуществляет совокупность радиальных мышц, называемых цилиарными мышцами. (Когда мы достигаем примерно 45 лет, хрусталик становится более жестким и мы постепенно утрачиваем способность фокусировать. Чтобы обойти это существенное возрастное неудобство, Бенджамин Франклин изобрел бифокальные очки.) Рефлекс, приводящий к сокращению цилиарных мышц и делающий хрусталик более выпуклым, определяется зрительным входом и тесно связан с рефлексом, контролирующим сопутствующий поворот глаз.
 Рис. 18. Офтальмолог, рассматривая глазное дно, видит нечто сходное с этим фотоснимком нормальной сетчатки. Сосок зрительного нерва расположен слева; здесь в сетчатку входят артерии и из нее выходят (более темные) вены. Темно-красная зона у самого края справа — макула; в центре этой области располагается центральная ямка, на снимке не показанная. Темная зона вверху слева — нормальная меланиновая пигментация.
Рис. 18. Офтальмолог, рассматривая глазное дно, видит нечто сходное с этим фотоснимком нормальной сетчатки. Сосок зрительного нерва расположен слева; здесь в сетчатку входят артерии и из нее выходят (более темные) вены. Темно-красная зона у самого края справа — макула; в центре этой области располагается центральная ямка, на снимке не показанная. Темная зона вверху слева — нормальная меланиновая пигментация.
 Рис. 19. Глазное яблоко и мышцы, контролирующие его положение. Роговица и хрусталик фокусируют световые лучи на заднюю стенку глаза. Хрусталик регулирует фокусировку близких и удаленных объектов — его выпуклость соответственно увеличивается и уменьшается.
Рис. 19. Глазное яблоко и мышцы, контролирующие его положение. Роговица и хрусталик фокусируют световые лучи на заднюю стенку глаза. Хрусталик регулирует фокусировку близких и удаленных объектов — его выпуклость соответственно увеличивается и уменьшается.
Две другие совокупности мышечных волокон изменяют диаметр зрачка и таким образом регулируют количество света, поступающего в глаз, точно так же, как в фотографическом аппарате это делает диафрагма. Система радиальных волокон, напоминающих спицы колеса, расширяет зрачок; другие, кольцевые волокна сужают его. Наконец, самоочистка передней поверхности роговицы достигается морганием век и смазкой из слезных желез. Роговица обильно снабжена нервами, чувствительными к прикосновению и боли, — вот почему малейшее раздражение пылинками вызывает рефлекс, который ведет к морганию и усиленному выделению слез.
 Рис. 20. Свет входит в глаз через прозрачную роговицу, которая вносит наибольший вклад в преломление световых лучей. Белое пятно на зрачке — отражение света.
Рис. 20. Свет входит в глаз через прозрачную роговицу, которая вносит наибольший вклад в преломление световых лучей. Белое пятно на зрачке — отражение света.
Сетчатка
Вся сложная суперструктура, описанная выше, существует для того, чтобы могла работать сетчатка, которая сама является удивительной структурой. Она преобразует свет в нервные сигналы, позволяет нам видеть в условиях от звездной ночи до солнечного дня, различает длины волн, что дает нам возможность видеть цвета, и обеспечивает точность, достаточную, чтобы заметить человеческий волос или соринку с расстояния в несколько метров. Сетчатка — это часть мозга, отделившаяся от него на ранних стадиях развития, но все еще связанная с ним посредством пучка волокон — зрительного нерва. Подобно многим другим структурам центральной нервной системы, сетчатка имеет форму пластинки, в данном случае толщиной приблизительно в четверть миллиметра. Она состоит из трех слоев тел нервных клеток, разделенных двумя слоями синапсов, образованных аксонами и дендритами этих клеток. Слой клеток на задней поверхности сетчатки содержит светочувствительные рецепторы — палочки и колбочки. Палочки, значительно более многочисленные, чем колбочки, ответственны за наше зрение при слабом свете и отключаются при ярком освещении. Колбочки не реагируют на слабый свет, но ответственны за способность видеть тонкие детали и за цветовое зрение. Число палочек и колбочек заметно изменяется в разных частях сетчатки. В самом центре, где способность нашего зрения различать тонкие детали максимальна, имеются только колбочки. Эту лишенную палочек зону диаметром примерно полмиллиметра называют центральной ямкой. Колбочки имеются по всей сетчатке, но наиболее плотно упакованы в центральной ямке. Поскольку палочки и колбочки расположены на задней поверхности сетчатки, поступающий свет должен пройти через два других слоя, чтобы их стимулировать. Мы точно не знаем, почему сетчатка устроена таким странным образом — как бы перевернута. Одна из возможных причин — то, что позади рецепторов находится слой клеток, содержащих черный пигмент меланин (он имеется также в коже). Меланин поглощает прошедший через сетчатку свет, не давая ему отражаться назад и рассеиваться внутри глаза; он играет ту же роль, что и черная окраска внутренности фотокамеры. Клетки, содержащие меланин, способствуют также химическому восстановлению светочувствительного зрительного пигмента, который обесцвечивается на свету (см. гл. 8). Для выполнения обеих функций нужно, чтобы меланин находился поблизости от рецепторов. Если бы рецепторы лежали впереди, пигментные клетки должны были бы располагаться между ними и следующим слоем нервных клеток, в области, уже заполненной аксонами, дендритами и синапсами. Как бы то ни было, слои перед рецепторами довольно прозрачны и, вероятно, не сильно вредят четкости изображения. Однако на центральном миллиметре, где наше зрение наиболее остро, последствия даже небольшого уменьшения четкости были бы катастрофическими, и эволюция, видимо, «постаралась» смягчить их — сместила другие слои к периферии, образовав здесь кольцо из утолщенной сетчатки и обнажив центральные колбочки так, что они оказались на самой поверхности. Образующееся маленькое углубление и есть центральная ямка. Двигаясь от заднего слоя к переднему, мы попадаем в средний слой сетчатки, расположенный между палочками и колбочками, с одной стороны, и ганглиозными клетками — с другой. Этот слой содержит нейроны трех типов: биполярные, горизонтальные и амакриновые клетки. Биполярные клетки имеют входы от рецепторов, как показано на рис. 21, и многие из них передают сигналы непосредственно ганглиозным клеткам. Горизонтальные клетки соединяют рецепторы и биполярные клетки сравнительно длинными связями, идущими параллельно сетчаточным слоям; сходным образом амакриновые клетки связывают биполярные клетки с ганглиозными.
 Рис. 21. В увеличенном фрагменте сетчатки справа показано относительное расположение трех ее слоев. Это покажется удивительным, но прежде чем свет достигнет палочек и колбочек, он должен пройти через слои ганглиозных и биполярных клеток.
Рис. 21. В увеличенном фрагменте сетчатки справа показано относительное расположение трех ее слоев. Это покажется удивительным, но прежде чем свет достигнет палочек и колбочек, он должен пройти через слои ганглиозных и биполярных клеток.
Слой нейронов на передней стороне сетчатки содержит ганглиозные клетки, аксоны которых проходят по поверхности сетчатки, собираются в пучок у слепого пятна и покидают глаз, образуя зрительный нерв. В каждом глазу около 125 миллионов палочек и колбочек, но всего 1 миллион ганглиозных клеток. Ввиду такого различия возникает вопрос: каким образом может сохраняться детальная зрительная информация? Изучение связей между клетками сетчатки может помочь разрешить эту проблему. Можно представить себе два пути информационного потока через сетчатку: прямой путь, идущий от фоторецепторов к биполярным и далее к ганглиозным клеткам, и непрямой путь, при котором между рецепторами и биполярами могут быть включены еще горизонтальные клетки, а между биполярами и ганглиозными клетками — амакриновые клетки (см. рис. 22, иллюстрирующий эти прямые и непрямые связи). Эти связи уже были весьма обстоятельно изучены Рамон-и-Кахалом примерно в 1900 году. Прямой путь весьма специфичен, или компактен, в том смысле, что одна биполярная клетка имеет входы лишь от одного рецептора или от сравнительно небольшого их числа, а одна ганглиозная клетка — от одного или сравнительно немногих биполяров. Непрямой путь более диффузен, или «размыт», благодаря более широким боковым связям. Общая площадь, занятая рецепторами, связанными с одной ганглиозной клеткой по прямому и непрямому путям, составляет всего около миллиметра. Эта зона, как вы, возможно, помните из главы 1, является рецептивным полем ганглиозной клетки — областью сетчатки, световая стимуляция которой может влиять на импульсацию данной ганглиозной клетки.
 Рис. 22. Поперечный разрез сетчатки приблизительно посредине между центральной ямкой и дальней периферией, где палочек больше, чем колбочек. Полной высоте рисунка в натуре соответствует примерно четверть миллиметра.
Рис. 22. Поперечный разрез сетчатки приблизительно посредине между центральной ямкой и дальней периферией, где палочек больше, чем колбочек. Полной высоте рисунка в натуре соответствует примерно четверть миллиметра.
Эта общая схема верна для всей сетчатки, но в деталях связей имеются большие различия между центральной ямкой, куда проецируется направление взора и где наша способность видеть тонкие детали максимальна, и периферией сетчатки, где острота зрения резко снижается. При переходе от центральной ямки к периферии сеть прямых путей от рецепторов к ганглиозным клеткам становится совершенно иной. В центральной ямке или около нее на прямом пути, как правило, одна колбочка связана с одной биполярной клеткой, а один биполяр — с одной ганглиозной клеткой. Однако по мере постепенного перехода к внешним областям все больше рецепторов конвергируют на биполярах, а биполяров — на ганглиозных клетках. Эта высокая степень конвергенции, которую мы видим в большей части сетчатки, вместе с весьма компактными путями в самом центре и около него позволяют понять, почему, несмотря на отношение 125:1 между числом рецепторов и числом волокон зрительного нерва, некоторая часть сетчатки (ее центр) может все-таки обеспечивать острое зрение. Общая схема сетчаточных путей с их прямым и непрямым компонентами была известна многие годы, а их связь с остротой зрения поняли задолго до того, как удалось выяснить роль непрямого пути. Ее понимание внезапно стало возможным, когда начали изучать физиологию ганглиозных клеток.
Рецептивные поля ганглиозных клеток сетчатки: выход глаза
При изучении сетчатки мы сталкиваемся с двумя главными проблемами. Во-первых, каким образом палочки и колбочки преобразуют поглощаемый ими свет в электрические и химические сигналы? Во-вторых, как последующие клетки двух других слоев — биполяры, горизонтальные, амакриновые и ганглиозные — интерпретируют эту информацию? Прежде чем обсуждать физиологию рецепторов и промежуточных клеток, я хочу забежать вперед и описать выходные сигналы сетчатки, представленные активностью ганглиозных клеток. Содержательной, удобной и компактной характеристикой нейрона, а тем самым и информации, доставляемой его выходными сигналами, может служить карта его рецептивного поля. Она может помочь нам понять, почему клетки промежуточных уровней соединены именно так, а не иначе, и объяснить назначение прямого и непрямого путей. Если мы узнаем, что́ ганглиозные клетки сообщают мозгу, мы значительно продвинемся в понимании работы сетчатки в целом. Примерно в 1950 году Стивен Куффлер впервые зарегистрировал реакции ганглиозных клеток сетчатки на световые пятна у млекопитающего, а именно у кошки. Он работал тогда в Уилмеровском офтальмологическом институте при больнице Джонса Гопкинса. Ретроспективно можно сказать, что выбор животного оказался удачным, поскольку сетчатка кошки как будто бы не обладает ни сложностью реакций на движение, наблюдаемой у лягушки или кролика, ни особенностями, связанными с цветом, как у рыб, птиц или обезьян.
 Рис. 23. Стивен Куффлер на лабораторном пикнике. Снимок сделан около 1965 года.
Рис. 23. Стивен Куффлер на лабораторном пикнике. Снимок сделан около 1965 года.
Куффлер использовал световой стимулятор, сконструированный С. Талботом. С помощью этого оптического прибора — видоизмененного медицинского офтальмоскопа — можно было равномерно освещать всю сетчатку постоянным слабым фоновым светом, а также проецировать маленькие, более яркие пятнышки, непосредственно наблюдая как стимул, так и кончик электрода. Фоновый свет позволял стимулировать либо палочки, либо колбочки, либо рецепторы обоих типов, поскольку при очень ярком освещении работают только колбочки, а при слабом — только палочки. Куффлер отводил реакции внеклеточными электродами, вводимыми через склеру (белая часть глаза) непосредственно в сетчатку с передней ее стороны. Находить ганглиозные клетки было нетрудно, так как они лежат под самой поверхностью сетчатки и довольно крупные. При постоянном рассеянном фоновом свете и даже в абсолютной темноте большинство ганглиозных клеток сетчатки проявляет стационарную, несколько нерегулярную активность с частотой от 1–2 до примерно 20 импульсов в секунду. Так как можно было бы ожидать, что в полной темноте клетки должны молчать, эта импульсация сама по себе оказалась неожиданной. Применяя маленькое световое пятнышко, Куффлер смог отыскивать на сетчатке области, с которых он мог влиять на импульсацию ганглиозных клеток — увеличивать ее или подавлять. Такие области и были рецептивными полями соответствующих ганглиозных клеток. Как и следовало ожидать, рецептивное поле обычно окружало кончик электрода или находилось очень близко к нему. Вскоре выяснилось, что ганглиозные клетки бывают двух типов, и по причинам, которые я скоро объясню, Куффлер назвал их клетками с on-центром и клетками с off-центром. Клетка с on-центром разряжается с заметно повышенной частотой, если небольшое пятнышко света появляется где-то внутри определенной зоны в центре или около центра рецептивного поля. Если слушать разряды такой клетки через громкоговоритель, то сначала вы услышите спонтанную импульсацию, отдельные случайные щелчки, а затем, после включения света, возникает залп импульсов, напоминающий пулеметную очередь. Мы назовем эту форму реакции on-реакцией. Когда Куффлер перемещал световое пятнышко чуть подальше от центра рецептивного поля, свет подавлял спонтанную импульсацию клетки, а при выключении света клетка давала залп учащенных импульсов, длившийся около секунды. Такую последовательность — подавление импульсации во время воздействия света и разряд после его выключения — мы называем off-реакцией. Исследование рецептивного поля этого типа вскоре показало, что оно четко подразделено на круглую on-зону и окаймляющую ее намного бо́льшую кольцеообразную off-зону. Чем бо́льшая часть данной зоны, on- или off-, заполнялась стимулом, тем сильнее был ответ, так что максимальные on-реакции получались на круглое пятно определенного диаметра, а максимальные off-реакции — на кольцо определенных размеров (с определенным внутренним и наружным диаметром). На рис. 24 приведены типичные записи реакций на такие стимулы. Центральная и периферическая зоны проявляли взаимный антагонизм: реакция на пятно в центре уменьшалась в результате вспышки второго пятна на периферии, как если бы клетку заставляли разряжаться одновременно быстрее и медленнее. Наиболее впечатляющая демонстрация этого взаимодействия между центром и периферией получалась при покрытии всего рецептивного поля одним большим пятном. Это вызывало намного более слабую реакцию, чем при заполнении пятном только центра; для некоторых же клеток эффекты стимуляции обеих зон полностью погашали друг друга. Прямо противоположным было поведение клетки с off-центром. Ее рецептивное поле состояло из маленького центра, с которого получалась off-реакция, и периферии, дававшей on-реакцию. Клетки обоих типов были перемешаны и встречались примерно одинаково часто. Клетки с off-центром разряжаются с наибольшей частотой в ответ на черное пятно на белом фоне, поскольку при этом освещается только периферия ее рецептивного поля. В природе темные объекты, по-видимому, столь же распространены, что и светлые; этим можно объяснить, почему информация от сетчатки передается клетками как с on-центром, так и с off-центром.
 Рис. 24. Слева: четыре записи ответов типичной ганглиозной клетки с on-центром. Каждая запись получена при одиночной развертке луча осциллографа длительностью 2,5 секунды. Из-за столь медленной развертки восходящая и нисходящая фазы импульса сливаются, так что каждый импульс имеет вид одной вертикальной линии. Слева показаны стимулы. Верхняя запись — состояние покоя (стимула нет): импульсы возникают редко и более или менее случайно. Три нижние записи — реакции на небольшое (оптимальной величины) пятно, на крупное пятно, покрывающее центр и периферию рецептивного поля, и на кольцо, покрывающее только периферию. Справа: реакции ганглиозной клетки с off-центром на такой же набор стимулов.
Рис. 24. Слева: четыре записи ответов типичной ганглиозной клетки с on-центром. Каждая запись получена при одиночной развертке луча осциллографа длительностью 2,5 секунды. Из-за столь медленной развертки восходящая и нисходящая фазы импульса сливаются, так что каждый импульс имеет вид одной вертикальной линии. Слева показаны стимулы. Верхняя запись — состояние покоя (стимула нет): импульсы возникают редко и более или менее случайно. Три нижние записи — реакции на небольшое (оптимальной величины) пятно, на крупное пятно, покрывающее центр и периферию рецептивного поля, и на кольцо, покрывающее только периферию. Справа: реакции ганглиозной клетки с off-центром на такой же набор стимулов.
Если постепенно увеличивать пятно, реакция возрастает, пока не заполнится центр рецептивного поля, а затем она начинает убывать, по мере того как захватывается все бо́льшая и бо́льшая доля периферии, как это можно видеть на приведенном графике (рис. 26). При пятне, покрывающем поле целиком, либо слегка преобладает действие центра, либо реакция нулевая. Это позволяет понять, почему нейрофизиологи до Куффлера были столь неудачливыми: при регистрации активности от ганглиозных клеток они всегда использовали рассеянный свет — далеко не лучший стимул. Можно представить себе удивление исследователей, когда магниевая вспышка, направленная прямо в глаз животного, вызывала столь слабые реакции или не вызывала их вовсе. Казалось бы, следовало ожидать, что освещение всех рецепторов, гарантируемое при такой вспышке, будет наиболее сильным, а не слабым стимулом. Ошибка здесь — в забвении того, насколько важны для нервной системы тормозные синапсы. Не располагая ничем, кроме схемы связей вроде изображенной на рис. 14, мы не можем предсказать влияние данного стимула на любую данную клетку, если не знаем, какие синапсы возбуждающие, а какие тормозные. В начале 50-х годов, когда Куффлер регистрировал реакции ганглиозных клеток, важность торможения в нервной системе только начинали осознавать.
 Рис. 25. Два главных типа рецептивных полей ганглиозных клеток сетчатки — с on-центром и тормозящей периферией и с off-центром и возбуждающей периферией. Знак «плюс» — область, дающая on-реакции; знак «минус» — область, дающая off-реакции.
Рис. 25. Два главных типа рецептивных полей ганглиозных клеток сетчатки — с on-центром и тормозящей периферией и с off-центром и возбуждающей периферией. Знак «плюс» — область, дающая on-реакции; знак «минус» — область, дающая off-реакции.
 Рис. 26. Если стимулировать одиночную ганглиозную клетку с on-центром все более крупными световыми пятнами, реакция будет постепенно усиливаться вплоть до пятна с величиной около 1 градуса. Это совпадает с величиной центра. Дальнейшее увеличение пятна ведет к уменьшению реакции, так как при этом пятно начинает захватывать антагонистическую периферию. При размерах пятна более 3 градусов реакция перестает уменьшаться, поэтому 3 градуса — это поперечник всего рецептивного поля, включая центр и периферию.
Рис. 26. Если стимулировать одиночную ганглиозную клетку с on-центром все более крупными световыми пятнами, реакция будет постепенно усиливаться вплоть до пятна с величиной около 1 градуса. Это совпадает с величиной центра. Дальнейшее увеличение пятна ведет к уменьшению реакции, так как при этом пятно начинает захватывать антагонистическую периферию. При размерах пятна более 3 градусов реакция перестает уменьшаться, поэтому 3 градуса — это поперечник всего рецептивного поля, включая центр и периферию.
Прежде чем перейти к описанию рецепторов и других клеток сетчатки, я хочу рассмотреть еще три вопроса относительно рецептивных полей. Первый из них касается общего понятия «рецептивное поле», а два других — некоторых особенностей рецептивных полей ганглиозных клеток сетчатки: их перекрывания и их размеров.
Понятие рецептивного поля
Термин рецептивное поле в узком смысле означает просто совокупность рецепторов, посылающих данному нейрону сигналы через один или большее число синапсов. В зрительной системе это всего лишь некоторая область сетчатки, но со времен Куффлера и благодаря его работам этот термин постепенно стал использоваться в значительно более широком смысле. Ганглиозные клетки сетчатки исторически были первым примером нейронов, рецептивные поля которых обладают внутренней структурой: стимуляция разных частей рецептивного поля дает качественно различные реакции, а стимуляция значительной части поля может приводить к взаимному погашению эффектов от отдельных частей, а не к их сложению. Характеризуя рецептивное поле, в настоящее время обычно описывают его субструктуру, т.е., иными словами, указывают, как нужно стимулировать ту или другую его зону, чтобы вызвать реакцию клетки. Когда мы говорим о «картировании рецептивного поля клетки», мы часто подразумеваем не просто очерчивание его границ на сетчатке или на экране, стоящем перед животным, но также и описание его субструктуры. По мере дальнейшего продвижения в глубь центральной нервной системы, где рецептивные поля нейронов становятся все сложнее, соответственно будет возрастать и сложность их описаний. Карты рецептивных полей особенно полезны тем, что позволяют предсказывать поведение клетки. Предположим, например, что в ганглиозном слое сетчатки мы стимулируем клетку с on-центром с помощью светового прямоугольника, ширина которого точно соответствует центру рецептивного поля, а длина больше диаметра всего поля вместе с периферией. По карте для клетки с on-центром, изображенной на рис. 25, мы могли бы предсказать, что такой стимул вызовет сильную реакцию, так как он покрывает весь центр и лишь небольшую долю антагонистического окружения. Кроме того, основываясь на радиальной симметрии карты, мы сможем предсказать, что величина реакции клетки не будет зависеть от ориентации световой полосы. Оба предсказания подтверждаются в опыте.
Перекрывание рецептивных полей
С перекрыванием рецептивных полей связан важный вопрос о том, что же делает в ответ на световой стимул некоторая популяция клеток, например выходных клеток сетчатки. Чтобы понять, что делают ганглиозные или любые другие клетки сенсорной системы, мы должны использовать два подхода к проблеме. Картируя рецептивное поле, мы задаемся вопросом, какой нужен стимул, чтобы вызвать реакцию одной клетки. Но мы также хотим знать, как тот или иной конкретный сетчаточный стимул влияет на всю популяцию ганглиозных клеток. Для ответа на второй вопрос следует сначала выяснить, что общего между собой имеют две соседние ганглиозные клетки, расположенные в сетчатке бок о бок. Данное выше описание рецептивных полей ганглиозных клеток может ввести в заблуждение, если представлять их себе в виде мозаики неперекрывающихся маленьких кружочков на сетчатке, чем-то вроде кафеля, которым выложен пол ванной комнаты. На самом деле соседние ганглиозные клетки получают входные сигналы от сильно перекрывающихся и обычно лишь незначительно разнящихся групп рецепторов (т.е. рецептивных полей), как это схематически показано на рис. 27.
 Рис. 27. Рецептивные поля двух соседних ганглиозных клеток сетчатки обычно перекрываются. Наименьшее световое пятнышко, которое мы можем проецировать на сетчатку, оказывает, по-видимому, влияние на сотни ганглиозных клеток, из которых одни имеют off-центры, а часть — on-центры; оно попадает на центры одних рецептивных полей и на периферию других.
Рис. 27. Рецептивные поля двух соседних ганглиозных клеток сетчатки обычно перекрываются. Наименьшее световое пятнышко, которое мы можем проецировать на сетчатку, оказывает, по-видимому, влияние на сотни ганглиозных клеток, из которых одни имеют off-центры, а часть — on-центры; оно попадает на центры одних рецептивных полей и на периферию других.
Рассмотрев упрощенную схему на рис. 28, легко понять, чем это обусловлено: ганглиозные клетки, окрашенные красным и голубым, имеют входы от перекрывающихся областей, соответственно окрашенных на поперечном сечении. Вследствие дивергенции, при которой на каждом уровне одна клетка образует синапсы со многими другими клетками, один рецептор может оказывать влияние на сотни или тысячи ганглиозных клеток. Он будет находиться в центрах рецептивных полей одних клеток и на периферии полей других клеток. Этот рецептор будет возбуждать некоторые нейроны через их центры, если это клетки с on-центром, или через их периферию, если это клетки с off-центром; и он будет одновременно тормозить другие нейроны через их центры или периферию. Таким образом, маленькое световое пятнышко, появившееся на сетчатке, может вызвать разнообразную активность многих клеток.
 Рис. 28. Две соседние ганглиозные клетки получают по прямому пути входные сигналы от двух перекрывающихся групп рецепторов. Области сетчатки, занятые этими рецепторами, составляют центры их рецептивных полей, представленные в плане большими перекрывающимися кругами.
Рис. 28. Две соседние ганглиозные клетки получают по прямому пути входные сигналы от двух перекрывающихся групп рецепторов. Области сетчатки, занятые этими рецепторами, составляют центры их рецептивных полей, представленные в плане большими перекрывающимися кругами.
Размеры рецептивных полей
Попробуем теперь соотнести события, развертывающиеся на сетчатке, с повседневным зрительным восприятием внешнего мира. У разных ганглиозных клеток рецептивные поля различны по величине. Особенно заметным и регулярным образом изменяются размеры центров рецептивных полей: они минимальны в центральной ямке сетчатки, где острота зрения, или способность различать небольшие объекты, выше всего; чем дальше от центральной ямки, тем поля становятся все крупнее, а острота зрения соответственно понижается. Величину рецептивного поля можно измерять двумя способами. Проще всего указывать его размеры непосредственно на сетчатке. Недостаток такого способа в том, что с обыденной точки зрения он не будет достаточно осмысленным. Вместо этого можно измерять величину проекции рецептивного поля на внешний мир, например его диаметр на экране, перед которым находится животное; при этом, однако, нужно указывать, как далеко от глаз расположен экран. Во избежание этих трудностей лучше, пожалуй, выражать величину рецептивного поля углом, под которым само животное будет видеть проекцию поля на экран, как показано на рис. 29. Мы рассчитываем этот угол в радианах путем простого деления диаметра поля на расстояние до экрана, но я буду пользоваться градусами путем пересчета по формуле: (радианы × π)/180. Один миллиметр на сетчатке человека примерно соответствует углу 3,5 градуса. При расстоянии до экрана 135 см одному градусу соответствуют на экране 2,5 см. Луна и Солнце, видимые с Земли, имеют почти одинаковые размеры в полградуса.
 Рис. 29. Одному миллиметру на сетчатке соответствует угол зрения в 3,5 градуса. Таким образом, на экране, удаленном на расстояние 1,5 метра, 1 миллиметр на сетчатке соответствует примерно 89 миллиметрам.
Рис. 29. Одному миллиметру на сетчатке соответствует угол зрения в 3,5 градуса. Таким образом, на экране, удаленном на расстояние 1,5 метра, 1 миллиметр на сетчатке соответствует примерно 89 миллиметрам.
У обезьян наименьшие из измеренных до сих пор центров рецептивных полей имеют диаметр около 2 угловых минут, или 10 мкм (0,01 миллиметра) на сетчатке. Эти ганглиозные клетки расположены, по-видимому, вне центральной ямки, но поблизости от нее. В самой ямке диаметр колбочек и расстояния между их центрами составляют около 2,5 мкм; это хорошо согласуется с данными об остроте нашего зрения — мы способны различить две точки при видимом расстоянии между ними всего лишь 0,5 угловых минут. Кружок диаметром 2,5 мкм на сетчатке (угол 0,5 минуты) соответствует 25-центовой монетке, видимой с расстояния около 150 метров. На дальней периферии сетчатки центры рецептивных полей состоят из тысяч рецепторов и могут иметь диаметр в 1 градус и более. Таким образом, по мере удаления от центральной ямки отмечается явно не случайное хорошо согласованное изменение трех величин: падает острота зрения, возрастает число рецепторов, участвующих в отдельных прямых путях (к биполярам и далее к ганглиозным клеткам), и увеличивается диаметр центров рецептивных полей. Это помогает нам понять значение прямых и непрямых путей от рецепторов к ганглиозным нейронам, так как дает веские основания для вывода, что центр рецептивного поля определяется прямым путем, а антагонистическая периферия — непрямым путем и что острота зрения лимитируется прямыми путями. Для получения дополнительных данных в пользу такого вывода нужны были сведения об активности других клеток сетчатки; об этом пойдет речь в следующих разделах.
Фоторецепторы
Прошло немало лет, прежде чем был достигнут существенный прогресс в физиологии рецепторов, биполяров, горизонтальных и амакриновых клеток. Тому было множество причин: пульсация сосудов постоянно мешала попыткам удерживать микроэлектрод в одиночной клетке или рядом с ней; рецепторы, биполяры и горизонтальные клетки не генерируют импульсов, поэтому регистрация намного меньших градуальных потенциалов требует применения внутриклеточных методик; трудно с уверенностью сказать, в клетке какого типа (или рядом с какой клеткой) находится электрод. Некоторые из этих затруднений можно преодолеть надлежащим выбором животного; например, сетчатки холоднокровных позвоночных способны выживать, будучи извлечены из глаза и погружены в солевой раствор, насыщенный кислородом, и при этом отсутствие кровообращения исключает пульсацию артерий; у протея (род крупных саламандр) очень большие клетки, их активность легко регистрировать; рыбы, лягушки, черепахи, кролики и кошки — все эти животные имеют свои преимущества при исследованиях того или иного типа, поэтому при изучении физиологии сетчатки использовались разные виды. Трудность при работе с таким большим числом видов состоит в том, что детали организации сетчатки могут заметно различаться у разных животных. Кроме того, наши представления о сетчатке приматов, реакции которой трудно регистрировать, до недавнего времени в значительной мере основывались на результатах, полученных на других видах. Однако по мере преодоления технических трудностей ускоряется и прогресс исследований на приматах. В последние годы изучение реакции палочек и колбочек на свет очень сильно продвинулось вперед, и появилось ощущение, что мы начинаем понимать, как они работают. Палочки и колбочки различаются во многих отношениях. Наиболее важно различие в их относительной чувствительности: палочки чувствительны к очень слабому свету, колбочки требуют намного более яркого освещения. Я уже описывал различия в их распределении по сетчатке, наиболее заметное из них — отсутствие палочек в центральной ямке. Они различны и по форме: палочки длинные и тонкие, а колбочки короткие и конусообразные. Как палочки, так и колбочки содержат светочувствительные пигменты. Во всех палочках пигмент один и тот же; колбочки делятся на три типа, каждый из них со своим особым зрительным пигментом. Эти четыре пигмента чувствительны к различным длинам световых волн, и в случае колбочек эти различия составляют основу цветового зрения. Под воздействием света в рецепторах происходит процесс, называемый выцветанием. В этом процессе молекула зрительного пигмента поглощает фотон — единичный квант видимого света — и при этом химически превращается в другое соединение, хуже поглощающее свет или, быть может, чувствительное к другим длинам волн. Практически у всех животных, от насекомых до человека, и даже у некоторых бактерий этот рецепторный пигмент состоит из белка, к которому присоединена небольшая молекула, близкая к витамину A; она и представляет собой химически трансформируемую светом часть. Благодаря главным образом работам Джорджа Уолда из Гарварда, проведенным в 50-х годах, нам теперь многое известно о химии выцветания и последующего восстановления зрительных пигментов.
 Рис. 30. Этот срез периферической части сетчатки обезьяны проходит через слой палочек и колбочек. Маленькие белые пятнышки — палочки; более крупные черные участки с белыми точками в центре — колбочки.
Рис. 30. Этот срез периферической части сетчатки обезьяны проходит через слой палочек и колбочек. Маленькие белые пятнышки — палочки; более крупные черные участки с белыми точками в центре — колбочки.
Большинство обычных сенсорных рецепторов — химических, температурных или механических — деполяризуется в ответ на соответствующий стимул, т.е. они реагируют на возбуждающий стимул так же, как обычные нейроны; деполяризация ведет к высвобождению медиатора из аксонных окончаний (часто, как и в случае зрительных рецепторов, это не приводит к возникновению импульсов, вероятно из-за очень малой длины аксона). У беспозвоночных, от усоногих раков до насекомых, световые рецепторы ведут себя таким же образом, и до 1964 года предполагалось, что аналогичный механизм — деполяризация под влиянием света — действует также в палочках и колбочках позвоночных. В 1964 году японскому нейрофизиологу Цунео Томита, работавшему в университете Кейо в Токио, впервые удалось ввести микроэлектрод в колбочки сетчатки рыбы и получить столь неожиданный результат, что у многих современников он вызывал вначале серьезные сомнения. В темноте потенциал на мембране колбочки оказался необычайно низким для нервной клетки: приблизительно 50 милливольт вместо обычных 70. При освещении колбочки этот потенциал возрастал — мембрана гиперполяризовалась — в противоположность тому, чего следовало бы ожидать. В темноте фоторецепторы позвоночных явно больше деполяризованы (имеют более низкий мембранный потенциал), чем обычные нервные клетки в состоянии покоя, и деполяризация вызывает непрерывное высвобождение медиатора из окончаний их аксонов — в точности так, как это происходит в обычных рецепторах при стимуляции. Свет, повышая потенциал на мембране рецепторной клетки (т.е. гиперполяризуя ее), уменьшает выделение медиатора. Таким образом, стимуляция, как это ни странно на первый взгляд, выключает рецепторы. Открытие Томита помогает нам объяснить, почему волокна зрительного нерва у позвоночных столь активны в темноте: спонтанную активность проявляют именно рецепторы; многие биполярные и ганглиозные клетки, вероятно, делают попросту то, что им диктуют рецепторные клетки. В последующие десятилетия главные задачи состояли в том, чтобы выяснить, как свет вызывает гиперполяризацию рецептора и в особенности каким образом выцветание всего одной молекулы зрительного пигмента под действием одного фотона может привести в палочке к измеримому изменению мембранного потенциала. В настоящее время оба процесса достаточно хорошо поняты. Гиперполяризация на свету вызывается перекрытием потока ионов. В темноте часть рецепторной мембраны более проницаема для ионов натрия, чем остальная мембрана. Поэтому ионы натрия непрерывно входят здесь в клетку, а где-то в другом месте ионы калия выходят наружу. Поток ионов в темноте, или темновой ток, открыли в 1970 году Уильям Хейгинс, Ричард Пенн и Шуко Йосиками в Национальном институте артрита и нарушений метаболизма в Бетезде. Он вызывает деполяризацию покоящегося рецептора и тем самым — его постоянную активность. В результате выцветания зрительного пигмента на свету поры для натрия закрываются, темновой ток уменьшается и степень деполяризации мембраны становится меньше, т.е. клетка гиперполяризуется. Ее активность (высвобождение ею медиатора) ослабевает.
 Рис. 31. Одиночная колбочка (слева) и две палочки с колбочкой (справа) были отпрепарированы и окрашены осмиевой кислотой. Тонкий отросток наверху каждой клетки — наружный сегмент, содержащий зрительный пигмент. Волокна внизу идут к не показанным здесь синаптическим областям.
Рис. 31. Одиночная колбочка (слева) и две палочки с колбочкой (справа) были отпрепарированы и окрашены осмиевой кислотой. Тонкий отросток наверху каждой клетки — наружный сегмент, содержащий зрительный пигмент. Волокна внизу идут к не показанным здесь синаптическим областям.
В настоящее время в результате работ Евгения Фесенко с сотрудниками в Академии наук в Москве, Дениса Бейлора в Стэнфордском университете, Кин-Вай Яу в Техасском университете и других мы намного ближе подошли к пониманию связи между выцветанием пигмента и закрытием натриевых пор. Например, очень трудно было представить себе, как выцветание единственной молекулы могло бы привести к закрытию миллионов пор, необходимому для наблюдаемых изменений потенциала. В настоящее время выяснилось, что поры в рецепторе открываются с помощью молекул вещества, называемого циклическим гуанозинмонофосфатом (цГМФ). Выцветание молекулы зрительного пигмента приводит к целому каскаду событий. Белковая часть обесцвеченной молекулы пигмента активирует большое число молекул фермента трансдуцина, а каждая из них в свою очередь инактивирует сотни молекул цГМФ, обычно участвующих в открытии пор. Так в результате выцветания одной молекулы пигмента закрываются миллионы пор. Все это позволяет объяснить ряд явлений, бывших ранее загадочными. Во-первых, давно известно, что человек, адаптировавшийся к полной темноте, способен увидеть такую слабую вспышку света, при которой ни один рецептор не может получить более одного фотона. Как показывают расчеты, для ощущения вспышки нужно, чтобы в короткий промежуток времени около шести близко расположенных палочек были стимулированы фотонами. Теперь становится понятно, как одиночный фотон может возбудить палочку и заставить ее генерировать сигнал достаточной силы. Во-вторых, мы теперь можем объяснить неспособность палочек реагировать на изменения освещенности, если свет уже достаточно ярок. По-видимому, чувствительность палочек столь высока, что при сильной освещенности, например при солнечном свете, все натриевые поры закрыты, и дальнейшее усиление света может не давать никакого дополнительного эффекта. Тогда мы говорим, что палочки насыщены. Быть может, спустя несколько лет студенты-биологи будут видеть во всей этой повести о рецепторах просто еще одну вещь, которую нужно выучить. Надеюсь, однако, что этого не произойдет. Чтобы полностью оценить ее значение, надо было потратить многие годы, гадая о том, каким образом могут работать рецепторы, а затем внезапно — меньше чем за десяток лет — в результате эффектных исследований решить эту проблему. Волнение по этому поводу еще не стихло.
Биполярные и горизонтальные клетки
Горизонтальные и биполярные клетки вместе с амакриновыми клетками расположены в среднем слое сетчатки. Биполярные клетки занимают в сетчатке стратегическую позицию, поскольку все сигналы, возникающие в рецепторах и поступающие к ганглиозным клеткам, должны пройти через них. Это означает, что они входят в состав как прямых, так и непрямых путей. В отличие от этого горизонтальные клетки входят только в непрямые пути. Как можно видеть на рис. 33, горизонтальные клетки встречаются намного реже биполярных, которые в целом преобладают в среднем слое.
 Рис. 32. Зарегистрировать реакцию клетки в нервной системе — это одна проблема; другая заключается в том, чтобы, сделав такую запись, точно знать, от клетки какого типа она получена. На этой микрофотографии показана одиночная биполярная клетка в сетчатке золотой рыбки, зарегистрированная в 1971 году Акимити Канеко, который тогда работал в Гарвардской медицинской школе. То, что это именно биполярная, а не амакриновая или горизонтальная клетка, было доказано инъекцией через микроэлектрод флуоресцентного красителя проционового желтого. Краситель распространяется по всей клетке, выявляя ее форму. На этом поперечном срезе рецепторы расположены сверху.
Рис. 32. Зарегистрировать реакцию клетки в нервной системе — это одна проблема; другая заключается в том, чтобы, сделав такую запись, точно знать, от клетки какого типа она получена. На этой микрофотографии показана одиночная биполярная клетка в сетчатке золотой рыбки, зарегистрированная в 1971 году Акимити Канеко, который тогда работал в Гарвардской медицинской школе. То, что это именно биполярная, а не амакриновая или горизонтальная клетка, было доказано инъекцией через микроэлектрод флуоресцентного красителя проционового желтого. Краситель распространяется по всей клетке, выявляя ее форму. На этом поперечном срезе рецепторы расположены сверху.
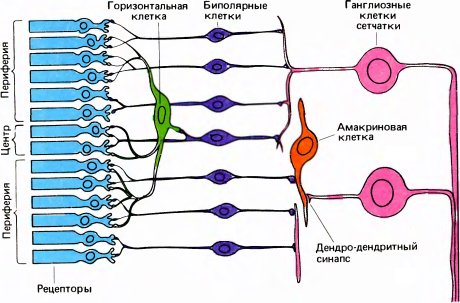 Рис. 33. Эта гипотетическая схема показывает, как могли бы создаваться рецептивные поля с центром и периферией. Центр, в данном случае off-центр, образует небольшая группа рецепторов, имеющих мощные возбуждающие синапсы с одиночной биполярной клеткой. Одна или несколько таких клеток соединены с ганглиозной клеткой, формируя ее центр. Периферию рецептивного поля биполярной клетки создает значительно большее число рецепторов (включая те, которые принадлежат к центральной группе), соединенных с горизонтальной клеткой возбуждающими синапсами. Горизонтальная клетка может контактировать с биполярной клеткой или посылать возвратные сигналы рецепторам.
Если биполярная клетка имеет off-центр, синапсы на этой клетке от центральной группы рецепторов предположительно должны быть возбуждающими (поскольку рецепторы выключаются светом). Горизонтальная клетка предположительно тормозит биполярную клетку или сами рецепторы. Отметим два входных пути к ганглиозным клеткам — один непосредственно от биполяров, а другой — от биполяра через амакриновую клетку к ганглиозной.
Рис. 33. Эта гипотетическая схема показывает, как могли бы создаваться рецептивные поля с центром и периферией. Центр, в данном случае off-центр, образует небольшая группа рецепторов, имеющих мощные возбуждающие синапсы с одиночной биполярной клеткой. Одна или несколько таких клеток соединены с ганглиозной клеткой, формируя ее центр. Периферию рецептивного поля биполярной клетки создает значительно большее число рецепторов (включая те, которые принадлежат к центральной группе), соединенных с горизонтальной клеткой возбуждающими синапсами. Горизонтальная клетка может контактировать с биполярной клеткой или посылать возвратные сигналы рецепторам.
Если биполярная клетка имеет off-центр, синапсы на этой клетке от центральной группы рецепторов предположительно должны быть возбуждающими (поскольку рецепторы выключаются светом). Горизонтальная клетка предположительно тормозит биполярную клетку или сами рецепторы. Отметим два входных пути к ганглиозным клеткам — один непосредственно от биполяров, а другой — от биполяра через амакриновую клетку к ганглиозной.
До того как удалось зарегистрировать активность биполярных клеток, никто не знал, обладают ли их рецептивные поля, как у ганглиозных клеток, центром и периферией и делятся ли эти поля на два типа — с on-центром и с off-центром. Утвердительный ответ на эти вопросы, почти несомненно, означал бы, что открытая Куффлером организация ганглиозных клеток пассивно отражает организацию биполяров. Данные о том, что рецептивные поля биполярных клеток действительно имеют центр и периферию и представлены двумя типами, были впервые получены путем внутриклеточной регистрации активности Джоном Даулингом и Фрэнком Верблином в Гарвардских биологических лабораториях и Акимити Канеко в Гарвардской медицинской школе. Следующий вопрос — как устроены эти рецептивные поля. Для ответа на него мы должны начать с изучения связей между рецепторами, биполярами и горизонтальными клетками. Биполярная клетка посылает по направлению к рецепторам единственный дендрит. Он либо образует синапс с одним рецептором (всегда с колбочкой), либо расщепляется на веточки, синаптически контактирующие более чем с одним рецептором. Если с одним биполяром связаны два или несколько рецепторов, они совместно занимают сравнительно малый участок сетчатки. В любом случае эти рецепторы должны составлять центр рецептивного поля, поскольку занимаемая ими площадь соответствует центру поля по величине. Возникает еще один вопрос: каковы синапсы между рецепторами и биполярными клетками — возбуждающие они, тормозные или обоих типов? Биполярные клетки, подобно рецепторам и горизонтальным клеткам, не генерируют импульсов, но мы и здесь будем говорить об on-реакции, подразумевая деполяризацию в ответ на световой стимул и соответственно усиленное выделение медиатора в выходных синапсах, и об off-реакции, разумея гиперполяризацию и уменьшение выброса медиатора. Что касается входных синапсов, передающих биполярам сигналы от рецепторов, то у биполяров с off-центром они должны быть возбуждающими, поскольку сами рецепторы выключаются (гиперполяризуются) светом; у биполяров с on-центром входные синапсы должны быть тормозными. Чтобы понять, почему это так (если вас, как и меня самого, это путает), следует лишь представить себе влияния малого светового пятна. Рецепторы активны в темноте, а свет, вызывая гиперполяризацию, уменьшает их активность. Если синапс возбуждающий, биполяр будет активироваться в темноте, а инактивироваться насвету; если же синапс тормозный, биполяр в темноте тормозится, а свет, выключая рецептор, снимает это торможение, т.е. биполярная клетка активируется (это действительно нелегко сразу понять). Является ли рецепторно-биполярный синапс возбуждающим или тормозным, зависит либо от выделяемого рецептором медиатора, либо от типа каналов в постсинаптической мембране биполярной клетки. В настоящее время никто не думает, что один рецептор высвобождает два медиатора, и имеется много данных в пользу того, что биполяры двух типов имеют различные рецепторные молекулы. Прежде чем обсуждать, как создается периферия рецептивного поля биполярного нейрона, мы должны рассмотреть горизонтальные клетки. Горизонтальные клетки важны потому, что они, видимо, по меньшей мере частично ответственны за периферию рецептивных полей ганглиозных клеток сетчатки; они составляют ту часть непрямого пути, о которой мы знаем больше всего. Это крупные клетки, и они принадлежат к числу самых удивительных в нервной системе. Их отростки тесно контактируют с окончаниями многих фоторецепторов, которые распределены по площади, размеры которой велики по сравнению с участком, непосредственно связанным с одиночной биполярной клеткой. Каждый рецептор контактирует с обоими типами клеток второго порядка — биполярными и горизонтальными. Имеется несколько подтипов горизонтальных клеток, которые могут сильно различаться у разных животных; их самое необычное свойство, общее с амакриновыми клетками, — это отсутствие чего-либо похожего на типичный аксон. Прочитав несколько упрощенное описание нервных клеток в предыдущей главе, вы с удивлением можете спросить: каким образом нерв без аксона будет передавать информацию другим нейронам? Когда начали использовать в нейроанатомии электронный микроскоп, вскоре выяснилось, что в некоторых случаях дендриты могут быть пресинаптическими отростками и образуют синапсы на других нейронах, обычно на их дендритах. (И наоборот, иногда окончания аксонов могут быть постсинаптическими структурами по отношению к другим аксонам, которые на них оканчиваются. Отростки, отходящие от тел горизонтальных и амакриновых клеток, могут, по-видимому, выполнять функции как аксонов, так и дендритов. Своеобразны и синапсы, образуемые горизонтальными клетками с рецепторами: у них отсутствуют электронно-микроскопические признаки, обычно указывающие, в каком направлении передаются сигналы. Ясно, что рецепторы доставляют информацию горизонтальным клеткам через возбуждающие синапсы, поскольку в большинстве случаев горизонтальные клетки, подобно рецепторам, гиперполяризуются (т.е. выключаются) светом. Менее ясно, куда эти клетки посылают свои выходные сигналы. Известно, что у некоторых животных, например у черепах, они передают информацию обратно рецепторам; у других видов они образуют синапсы с дендритами биполярных клеток и, несомненно, передают им информацию; о приматах мы не располагаем какими-либо сведениями. Итак, горизонтальные клетки получают входные сигналы от рецепторов; их выход пока точно не известен, но он направлен либо к рецепторам, либо к биполярным клеткам, либо к тем и другим. То, что рецепторы передают горизонтальным клеткам информацию с относительно обширного участка сетчатки, означает, что рецептивные поля этих клеток должны быть большими. Так оно и есть: их величина примерно соответствует размерам рецептивных полей биполярных или ганглиозных клеток, включая центр вместе с периферией. Они однородны, и освещение их в любом месте вызывает гиперполяризацию, которая тем сильнее, чем больше световое пятно. Многие данные указывают на то, что горизонтальные клетки ответственны за периферию рецептивных полей биполярных клеток; других кандидатов на эту роль, в сущности, нет, поскольку это единственные клетки, связанные с рецепторами на столь обширном пространстве. Там, где горизонтальные клетки непосредственно контактируют с биполярами, синапсы к on-биполярам должны быть возбуждающими (ввиду тормозящего влияния света на периферию), а к off-биполярам — тормозными. Если же влияние осуществляется через рецепторы, синапсы должны быть тормозными. Резюмируем сказанное выше. Биполярные клетки имеют рецептивные поля с центром и периферией. Реакция центра определяется прямым входом от небольшой группы рецепторов; периферию определяет непрямой путь от более обширной области рецепторов, связанных с горизонтальными клетками, которые, вероятно, в свою очередь передают сигналы биполярам. Непрямой путь, кроме того, мог бы быть результатом обратной связи от горизонтальных клеток, тормозящей рецепторы.
Амакриновые клетки
Эти клетки удивительно разнообразны по форме и используют необычайно большое число нейромедиаторов, которых может быть более двадцати. Все амакриновые клетки имеют ряд общих особенностей. Во-первых, тела их расположены в среднем слое сетчатки, а отростки — в синаптической зоне между этим слоем и ганглиозными клетками; во-вторых, они образуют связи и с биполярными, и с ганглиозными клетками и таким образом создают между теми и другими альтернативный, непрямой путь; и наконец, у них нет аксонов, но зато их дендриты способны к образованию пресинаптических окончаний на других клетках. Амакриновые клетки, вероятно, выполняют много различных функций, большей частью неизвестных. Один их тип, по-видимому, участвует в специфических реакциях на движущиеся объекты, обнаруженных в сетчатке лягушки и кролика; другой тип участвует в создании пути, связывающего ганглиозные клетки с теми биполярами, которые имеют входы от палочек. Нет данных о том, чтобы амакрины участвовали в организации центра и периферии рецептивных полей ганглиозных клеток, но нельзя полностью исключить такую возможность. Функции большинства их форм остаются невыясненными; следует, вероятно, прямо сказать, что анатомические сведения об амакриновых клетках в целом значительно богаче физиологических.
Связи между биполярными и ганглиозными клетками
Мы видели, что основные особенности рецептивных полей ганглиозных клеток проявляются уже у биполярных клеток. Это оставляет открытым вопрос о том, какие преобразования информации происходят между биполярами и ганглиозными клетками. Вряд ли тут совсем ничего не происходит, если судить по сложности синаптического слоя между средним слоем и слоем ганглиозных клеток, поскольку мы часто встречаем здесь конвергенцию на прямом пути от биполярных клеток к ганглиозным, а также включение амакриновых клеток, функция которых не вполне понятна. Вероятно, все синапсы между биполярными и ганглиозными клетками возбуждающие; это означает, что биполярные клетки с on-центром передают сигналы ганглиозным клеткам с on-центром, а биполяры с off-центром — ганглиозным клеткам с off-центром. Это упрощает схему связей: ведь могло быть так, например, что клетки с on-центром через тормозные синапсы воздействовали бы на клетки с off-центром. Ну что же, и на том спасибо! До 1976 года не было известно, различаются ли по форме клетки с on-центром и с off-центром, но в этом году Ральф Нелсон, Хельга Колб и Эдвард Фамильетти из Национальных институтов здоровья в Бетезде внутриклеточно зарегистрировали реакции ганглиозных клеток кошки, идентифицировав их как клетки с on- или off-центром, а затем через микроэлектрод ввели вещество, окрашивающее все дендритное дерево. Сравнив разветвления дендритов у клеток двух типов, они увидели четкое различие: две совокупности дендритов оканчивались на двух отдельных субслоях в пределах синаптической зоны между средним слоем и слоем ганглиозных клеток. Дендриты клеток с off-центром всегда оканчивались ближе к среднему слою сетчатки, а дендриты клеток с on-центром — дальше от него. К тому времени в другой работе уже было показано, что биполярные клетки двух классов, образующие с рецепторами синапсы разной формы, различаются также и расположением своих аксонных окончаний: у одних аксоны оканчиваются там, где кончаются дендриты ганглиозных клеток с on-центром, а у других — там, где кончаются дендриты клеток с off-центром. Таким образом, стало возможным реконструировать весь путь от рецепторов к ганглиозным клеткам для обеих систем — с on- и с off-центрами. В результате был установлен неожиданный факт: на прямом пути именно система с off-центрами имеет возбуждающие синапсы на каждом уровне — от рецепторов к биполярам и от биполяров к ганглиозным клеткам, тогда как в системе с on-центрами синапсы между рецепторами и биполярами тормозные. Разделение биполярных и ганглиозных клеток на категории с on- и с off-центром наверняка должно иметь свои корреляты в восприятии. Клетки с off-центром отвечают на темное пятно точно таким же образом, как клетки с on-центром — на светлое пятно. Если наличие двух совокупностей клеток, отвечающих на темные и на светлые пятна, кажется нам неожиданным, то это, возможно, результат совершенно справедливых утверждений физиков, что темнота — это отсутствие света. Но темнота представляется нам вполне реальной, и теперь мы, видимо, находим биологическую основу этой реальности. Черное для нас так же реально, как и белое. Шрифт книги, которую вы сейчас читаете, в конце концов тоже черный. Совершенно так же обстоит дело с теплом и холодом. Из курса физики мы с удивлением узнаем, что холод — это всего лишь отсутствие тепла, а между тем он кажется нам точно такой же реальностью, особенно если вы выросли, как это было со мной, в морозном Монреале. Наша интуиция подтверждается, когда мы узнаём, что в коже имеются температурные рецепторы двух типов: одни реагируют на повышение температуры, другие — на ее понижение. Поэтому с биологической точки зрения холод столь же реален, как и тепло. Во многих сенсорных системах используются оппонентные па́ры: тепло/холод, черное/белое, поворот головы налево/направо, а также, как мы увидим в главе 8, желтое/синее и красное/зеленое. Причина этого, вероятно, в характере нервной импульсации и ее изменений. В принципе можно представить себе нервы с частотой разрядов, поддерживаемой на некотором достаточно высоком уровне, скажем 100 в секунду, и потому способных в ответ на оппонентные стимулы либо снижать, либо повышать частоту импульсов — вниз до нуля, а вверх, например, до 500 в секунду. Но поскольку на импульсы расходуется метаболическая энергия (весь входящий в нерв натрий должен откачиваться наружу), то для нервных клеток в состоянии покоя более экономным, вероятно, будет молчание или импульсация с низкой частотой, а организму выгодно иметь для каждой данной модальности две отдельные группы клеток, одна из которых разряжается при уменьшении, а другая при увеличении силы стимула.
Значение рецептивных полей с центром и периферией
Зачем эволюции понадобилось создавать столь любопытные образования, как рецептивные поля с центральной и периферической зонами? Иными словами, какую пользу они приносят животному? Отвечать на столь глубокие вопросы всегда нелегко, но мы можем попытаться высказать ряд правдоподобных соображений. Сообщения, которые глаз посылает мозгу, могут быть очень мало связаны с абсолютной освещенностью сетчатки, так как ганглиозные клетки сетчатки плохо реагируют на изменения силы рассеянного света. О чем действительно сигнализирует клетка, так это о результате сравнения количества света, падающего на определенный участок сетчатки, с его средним количеством в ближайшем окружении. Мы можем проиллюстрировать это сравнение следующим экспериментом. Найдем сначала клетку с on-центром и картируем ее рецептивное поле. Затем, равномерно осветив экран слабым постоянным фоновым светом, начнем включать и выключать пятнышко, точно заполняющее центр поля, начиная со столь слабого света, что его еще нельзя увидеть, и постепенно повышая интенсивность. При некоторой яркости мы начнем обнаруживать реакцию; отметим, что именно при этой яркости мы и сами начнем видеть это пятнышко. Измерив интенсивность фона и пятнышка фотометром, мы выясним, что пятнышко приблизительно на 2 % ярче фона. Теперь повторим всю процедуру, начиная с фонового света в пять раз более яркого. Будем постепенно повышать интенсивность локального стимула. В какой-то момент мы снова начнем обнаруживать реакции, и опять это произойдет при такой яркости, когда мы сами будем едва замечать световое пятнышко на новом фоне. Измерив стимулирующий свет, мы найдем, что он тоже в пять раз ярче предыдущего, т.е. пятнышко снова на 2 % ярче фона. Вывод таков, что как для нас, так и для клетки существенна относительная освещенность пятнышка и фона. Неспособность клетки хорошо реагировать на что-либо кроме различий в локальной интенсивности может показаться странной, так как при рассматривании крупного равномерно освещенного пятна его внутренность представляется нам такой же яркой, как и его края. Ганглиозная клетка, если учесть ее физиологию, может передавать информацию только о границах пятна; внутренность его мы видим как однородную, поскольку ганглиозные клетки с полями, находящимися внутри пятна, не сообщают о локальных различиях в освещенности. Аргументация эта кажется достаточно убедительной, и все же мы испытываем некоторое сомнение: логика логикой, но ведь внутренность пятна видна все-таки абсолютно ясно! Мы вновь и вновь будем сталкиваться с этой проблемой в последующих главах, и нам придется признать, что нервная система часто работает по принципам, противоречащим нашей интуиции. Рассуждая рационально, однако, мы должны согласиться, что видеть большое пятно с помощью только тех клеток, поля которых лежат на его границах (не нуждаясь в участии остальных клеток с центрами, распределенными по всему пятну), — это более экономный способ: если вы инженер, то вы, вероятно, именно так сконструировали бы соответствующую машину. В таком случае и машина, я думаю, тоже «представляла бы себе» пятно освещенным равномерно. В одном отношении слабые реакции нейрона или их отсутствие при воздействии диффузного света не должны вызывать удивления. Каждый, кто пытался фотографировать без экспонометра, знает, насколько плохо мы можем судить об абсолютной интенсивности света. Считайте, что нам везет, если мы можем устанавливать диафрагму своего фотоаппарата с ошибкой не более чем вдвое; да и это достигается не прямой оценкой «на глаз», а лишь в результате большого опыта, позволяющего, например, отметить, что в момент съемки на небе легкая облачность и что мы находимся на открытом пространстве в тени за час до захода солнца. Однако при пространственных сравнениях, когда нужно сказать, какая из двух соседних областей ярче или темнее, мы, как и ганглиозные клетки, судим очень точно. Как уже говорилось, мы способны производить такое сравнение при различии всего в 2 % — в точности так, как это могут делать наиболее чувствительные ганглиозные клетки в сетчатке обезьяны. Наряду с эффективностью эта система обладает еще одним важным преимуществом. Большинство объектов мы видим благодаря отраженному свету таких источников, как солнце или электрическая лампочка. Несмотря на изменения освещенности, создаваемой этими источниками, наша зрительная система сохраняет удивительную способность к неизменному восприятию объектов. Ганглиозная клетка сетчатки работает именно так, что это становится возможным. Рассмотрим следующий пример: газета выглядит примерно одинаково — белая бумага, черные буквы, — читаем ли мы ее в полумраке комнаты или на пляже в солнечный день. Предположим, что в обоих случаях мы измерим входящий в наши глаза свет от белой бумаги и от одной из черных букв заголовка. Вот какие цифры я получил, когда перешел из закрытого помещения на солнце во внутренний дворик Гарвардской медицинской школы:
(пусто) // На открытом воздухе // В комнате Белая бумага // 120 // 6.0 Черная бумага // 12 // 0,6
Сами цифры не вызывают сомнений. Снаружи свет в 20 раз ярче, чем в комнате, а черные буквы отражают приблизительно десятую долю света, отражаемого белой бумагой. Но эти цифры, когда вы видите их впервые, тем не менее поражают, так как они показывают, что черная буква на открытом воздухе посылает в наши глаза вдвое больше света, чем белая бумага при комнатном освещении. Отсюда ясно, что восприятие черного и белого не определяется самим по себе количеством отражаемого объектом света. Существенно то, каково это количество в сравнении с окружающим фоном. Экран выключенного черно-белого телевизора в нормально освещенной комнате выглядит белым или серовато-белым. Инженеры создали электронные механизмы, которые могут сделать экран ярче, но не способны затемнить его, и независимо от того, как он выглядит в выключенном виде, никакая его часть никак не может послать после включения меньше света. Мы, однако, хорошо знаем, что телевизор может дать нам ощущение насыщенного черного цвета. Самый темный участок изображения посылает в наши глаза по меньшей мере столько же света, что и при выключенном телевизоре. Отсюда следует, что «черное» и «белое» — не просто физические понятия: это биологические термины, они отражают результат вычислений, производимых нашей сетчаткой и мозгом при обработке воспринимаемой картины. Как мы увидим в главе 8, все соображения, которые я высказал здесь о черном и белом, применимы и к цвету. Видимый цвет объекта определяется не только приходящим от него светом, но также — в столь же значительной степени, как в случае черного и белого, — и светом, приходящим от всего остального. В результате то, что мы видим, становится независимым не только от интенсивности света, но и от его спектрального состава. И опять-таки это способствует постоянству восприятия видимой картины несмотря на заметные различия в яркости и спектральном составе падающего на нее света.
Заключение
Выходные сигналы от глаза после двух или трех синапсов содержат намного более усложненную информацию, чем мозаичное представление мира, кодируемое палочками и колбочками. Что мне кажется особенно интересным, так это неожиданность новых результатов; до Куффлера никто не мог бы даже предположить, что существует что-либо вроде рецептивных полей с центром и периферией и что по зрительному нерву практически не передаются сведения о таком малосущественном параметре, как уровень диффузного освещения. Точно так же никому не удалось даже приблизительно догадаться, что же поступает на следующие уровни зрительной системы — в головной мозг. Именно эта непредсказуемость и делает мозг столь привлекательным объектом — наряду с остроумием его механизмов (после того как нам удается их раскрыть).
4. Первичная зрительная кора
После того как в 1952 году была опубликована первая статья Куффлера о рецептивных полях ганглиозных клеток сетчатки с центром и периферией, открылись пути дальнейшей работы. Чтобы объяснить обнаруженные свойства этих клеток, нужны были дополнительные исследования на уровне сетчатки. Однако требовались также данные о следующих уровнях зрительной системы, чтобы понять, как мозг интерпретирует информацию, доставляемую глазами. Решение этих двух задач было сопряжено с огромными трудностями. При изучении центральных механизмов понадобилось несколько лет, чтобы разработать методику длительной (порядка нескольких часов) регистрации активности одиночной клетки. Еще труднее было подбирать зрительные стимулы, влияющие на эту активность.Топографическое отображение
До того как стало возможным продолжение работ в этом направлении, уже имелись кое-какие сведения об участках мозга, связанных со зрительной функцией: достаточно хорошо были прослежены начальные этапы передачи зрительной информации (рис. 35). Мы знали, что волокна зрительного нерва образуют синапсы с клетками наружного коленчатого тела (НКТ) и что аксоны клеток НКТ оканчиваются в первичной зрительной коре. Было также ясно, что эти связи — от сетчатки к НКТ и от НКТ к коре — имеют топографическую организацию. Говоря о топографическом отображении, мы имеем в виду, что предшествующая структура проецируется на последующую упорядоченным образом: если идти вдоль какой-либо линии на сетчатке, то проекции последовательных точек этой линии в НКТ и в коре также образуют одну непрерывную линию. Таким образом, волокна зрительного нерва, выходящие из небольшого участка сетчатки, все будут направляться к какому-то небольшому участку НКТ, а все волокна от небольшой зоны НКТ придут в определенную зону зрительной коры. Такая организация связей не покажется удивительной, если мы вспомним упрощенную схему нервной системы на рис. 13: клетки группируются здесь в структуру, напоминающую стопку пластин, причем каждая клетка любой пластины получает входы от некоторой компактной группы клеток предыдущей пластины. В сетчатке последовательные слои клеток расположены наподобие игральных карт в колоде, так что нервные волокна могут кратчайшим путем проходить с одного уровня на следующий. Клетки наружных коленчатых тел удалены на некоторое расстояние от клеток сетчатки, точно так же как кора удалена от НКТ и находится в другой части мозга. Тем не менее характер соединений между клетками остается тем же — проекции отдельных зон на более высокие уровни организованы так, как если бы эти зоны точно накладывались друг на друга.
 Рис. 34. В зрительной коре обезьяны окраска по Гольджи выявляет лишь несколько пирамидных клеток — ничтожную долю общего их числа в срезе. В действительности высота представленного здесь прямоугольного участка составляет около миллиметра. На микрофотографию наложено изображение (в том же масштабе) типичного вольфрамового электрода, применяемого при внеклеточной регистрации активности нейронов.
Рис. 34. В зрительной коре обезьяны окраска по Гольджи выявляет лишь несколько пирамидных клеток — ничтожную долю общего их числа в срезе. В действительности высота представленного здесь прямоугольного участка составляет около миллиметра. На микрофотографию наложено изображение (в том же масштабе) типичного вольфрамового электрода, применяемого при внеклеточной регистрации активности нейронов.
 Рис. 35. Зрительные пути в мозгу человека, от глаз до первичной зрительной коры (вид снизу). Пусть входное изображение попадает на те половины обеих сетчаток, которые окрашены здесь в красный цвет (это их правые половины, поскольку мозг перевернут). На эти области проецируется противоположная половина пространства (левое зрительное поле); в конечном счете входная информация передается в правую половину мозга (пути ее передачи тоже окрашены в красный цвет). Это происходит потому, что примерно половина волокон, образующих зрительный нерв, переходит в хиазме на другую сторону, а другая половина остается на той же стороне. Таким образом, во-первых, каждое полушарие получает информацию от обоих глаз; во-вторых, каждое полушарие получает информацию о противоположной половине видимого мира.
Рис. 35. Зрительные пути в мозгу человека, от глаз до первичной зрительной коры (вид снизу). Пусть входное изображение попадает на те половины обеих сетчаток, которые окрашены здесь в красный цвет (это их правые половины, поскольку мозг перевернут). На эти области проецируется противоположная половина пространства (левое зрительное поле); в конечном счете входная информация передается в правую половину мозга (пути ее передачи тоже окрашены в красный цвет). Это происходит потому, что примерно половина волокон, образующих зрительный нерв, переходит в хиазме на другую сторону, а другая половина остается на той же стороне. Таким образом, во-первых, каждое полушарие получает информацию от обоих глаз; во-вторых, каждое полушарие получает информацию о противоположной половине видимого мира.
 Рис. 36. Зрительный нерв в том месте, где он выходит из глаза, прерывая слои сетчатки, показанные слева и справа. Ширина области, представленной на микрофотографии, около 2 мм. Свободная зона вверху рисунка — это внутренняя среда глаза. На срезе видны слои сетчатки (сверху вниз): волокна зрительного нерва (светлые), три окрашенных слоя клеток и черный слой, содержащий пигмент меланин.
Рис. 36. Зрительный нерв в том месте, где он выходит из глаза, прерывая слои сетчатки, показанные слева и справа. Ширина области, представленной на микрофотографии, около 2 мм. Свободная зона вверху рисунка — это внутренняя среда глаза. На срезе видны слои сетчатки (сверху вниз): волокна зрительного нерва (светлые), три окрашенных слоя клеток и черный слой, содержащий пигмент меланин.
На выходе из глазного яблока волокна зрительного нерва просто собираются в пучок. Дойдя до НКТ, они расходятся и образуют своими окончаниями топографически упорядоченную проекцию. (Удивительно то, что в зрительном нерве на пути от сетчатки к НКТ эти волокна почти полностью перепутываются, но в НКТ снова «находят свои места».) Точно так же и волокна, выходящие из НКТ, расходятся в виде широкой полосы, которая идет через весь мозг к затылку и оканчивается в первичной зрительной коре, где проекция снова оказывается упорядоченной. После того как эти пути, пройдя через первичную зрительную кору и образовав синапсы в различных ее слоях, выходят из этой области и достигают других корковых зон, они опять образуют топографически упорядоченную проекцию. Поскольку на каждом уровне имеет место конвергенция связей, рецептивные поля в целом постепенно становятся все ближе, поэтому чем дальше от сетчатки, тем более размытым будет отображение зрительно воспринимаемой картины. Еще одним важным и давним свидетельством в пользу топографической организации зрительных путей служат клинические наблюдения. Если поврежден определенный участок первичной зрительной коры, то развивается «локальная слепота», как если бы была разрушена соответствующая область сетчатки. Итак, зрительный мир систематически отображен в структурах наружных коленчатых тел и коры. Однако в 50-х годах было неясно, что может означать такое отображение. В то время было еще не очевидно, что мозг обрабатывает получаемую информацию, преобразуя ее так, чтобы привести к более удобному для использования виду. Полагали, что зрительная сцена просто передается в мозг, а уж его задача — осмыслить ее (или эта задача, быть может, решается вообще не мозгом, а разумом). Из последующих глав мы узнаем, что такая нейронная структура, как первичная зрительная кора, производит глубокое преобразование поступающей информации. Нам почти ничего не известно о том, что происходит на дальнейших этапах ее переработки. Поэтому можно было бы сказать, что мы продвинулись не слишком далеко. Однако знание того, что определенная часть коры использует в своей работе вполне понятные принципы, дает основание для оптимизма — позволяет предполагать, что и остальные зоны коры работают так же. Возможно, настанет день, когда нам вообще не придется употреблять слово «разум».
Ответы клеток наружного коленчатого тела
Волокна, идущие в мозг от каждого глаза, проходят через зрительную хиазму (от названия греческой буквы «хи» — χ). В хиазме примерно половина волокон каждого зрительного нерва переходит на противоположную сторону мозга по отношению к данному глазу, а другая половина остается на той же стороне мозга. Пройдя хиазму, волокна направляются в несколько разных пунктов. Некоторые из них идут в нейронные структуры, имеющие отношение к таким специфическим реакциям, как движения глаз и зрачковый рефлекс. Однако большая часть волокон оканчивается в двух наружных коленчатых телах (НКТ). По сравнению с корой и множеством других отделов мозга эти тела устроены сравнительно просто — все или почти все из примерно полутора миллионов клеток в каждом НКТ имеют непосредственные входы от волокон зрительного нерва, и большинство клеток (но не все) посылают свои аксоны в кору мозга. Отсюда следует, что пути, проходящие через НКТ в кору, имеют только одно синаптическое переключение. Однако было бы ошибкой считать НКТ просто передаточной станцией. Сюда входят не только волокна из зрительного нерва, но и волокна, приходящие обратно из тех участков коры, на которые проецируются НКТ, а также из ретикулярной формации ствола мозга, имеющей отношение к процессам внимания и общей активации (arousal). Некоторые клетки НКТ имеют короткие аксоны (меньше миллиметра длиной), они не выходят за пределы НКТ, а образуют синаптические контакты с другими нейронами НКТ. Несмотря на эти усложнения, одиночные клетки НКТ отвечают на световые стимулы примерно так же, как и ганглиозные клетки сетчатки, и у них сходная структура рецептивных полей с on- и off-центрами и сходные ответы на цветовые стимулы. Таким образом, если говорить о переработке зрительной информации, НКТ, по-видимому, не производит никаких значительных преобразований сигналов. Что же касается незрительных входов НКТ и локальных синаптических связей, то пока мы просто ничего не знаем об их роли.
Представительство правой и левой сторон в зрительном пути
Волокна зрительного нерва распределяются между двумя НКТ не совсем обычным и на первый взгляд даже странным способом. Волокна от левой половины сетчатки левого глаза идут в НКТ той же стороны мозга, в то время как волокна от левой половины сетчатки правого глаза переходят в хиазме на другую сторону и, таким образом, попадают в то же левое НКТ (это показано на рис. 35). Аналогичным образом волокна от правых половин обеих сетчаток тоже оканчиваются в одном — правом — полушарии. Поскольку хрусталик создает на сетчатке перевернутое изображение, световые лучи, исходящие из правой половины зрительной сцены, проецируются на левые половины обеих сетчаток и информация передается в левое полушарие. Термином зрительные поля обозначаются те участки окружающего мира или зрительной сцены, которые видны обоими глазами. Правое зрительное поле включает все точки, лежащие справа от вертикали, проходящей через ту точку, которую мы фиксируем взглядом (рис. 37). Важно отличать зрительные поля, т.е. то, что мы видим во внешнем мире, от рецептивного поля, которое соответствует тому участку внешнего мира, который «видит» одиночная клетка. Перефразируя то, что говорилось в начале этого раздела, можно сказать, что информация от правой половины поля зрения передается в левое полушарие.
 Рис. 37. Правое зрительное поле простирается вправо почти на 90°. В этом легко убедиться, если, быстро сгибая и разгибая палец, медленно смещать его по кругу вправо. Вверх поле зрения простирается на 60° или около того, вниз — примерно на 75°, а влево, по определению, доходит до вертикали, проходящей через точку, которую вы фиксируете взглядом.
Рис. 37. Правое зрительное поле простирается вправо почти на 90°. В этом легко убедиться, если, быстро сгибая и разгибая палец, медленно смещать его по кругу вправо. Вверх поле зрения простирается на 60° или около того, вниз — примерно на 75°, а влево, по определению, доходит до вертикали, проходящей через точку, которую вы фиксируете взглядом.
Аналогичным образом устроены и многие другие отделы мозга. Например, осязательные и болевые сигналы от правой половины тела приходят в левое полушарие, а двигательное управление мышцами правой стороны тела осуществляется левым полушарием. Обширный инсульт левой половины мозга приводит к параличу и потере чувствительности правой половины лица, правой ноги и правой руки, а также к потере речи. Но, что менее известно, такой инсульт обычно сопровождается утратой зрения в правой половине зрительного поля, причем для обоих глаз. Чтобы установить наличие такой слепоты, невропатолог просит больного встать перед ним, закрыть один глаз и смотреть другим на кончик носа врача. Затем он начинает обследовать зрительные поля больного, перемещая в разные точки пространства свою руку или специальный зонд. В случае левостороннего инсульта можно показать, что больной ничего не видит справа от точки фиксации. Например, если врач поднимает в руке зонд и держит его между собой и больным немного выше своей головы, то при медленном движении руки справа налево (с точки зрения больного) больной ничего не видит до тех пор, пока белый рукав халата врача не пересечет среднюю линию и внезапно не появится в поле зрения больного. Если проверить другой глаз, то результат будет точно таким же. Полная правосторонняя гомонимная гемианопсия (так называют неврологи подобное выпадение половины поля зрения!) точно разделит и область центральной ямки (центр взора): если смотреть на слово was, фиксируя взглядом середину буквы а, то не будет видно буквы s, и даже от буквы а останется лишь ее левая половина — интересное, но весьма огорчительное переживание. Такого рода тесты ясно показывают, что от каждого глаза сигналы передаются в оба полушария и что, наоборот, каждое полушарие мозга получает входные сигналы от обоих глаз. Это может показаться странным: после того что было сказано об осязательной и болевой чувствительности и о системе управления движениями, читатель может предположить, что левый глаз должен посылать сигналы в правое полушарие и наоборот. Однако в случае зрительной системы каждое полушарие мозга имеет дело не с противоположной стороной тела, а с противоположной половиной окружающего пространства. Но, впрочем, ситуация, когда сигналы от левого глаза передаются в основном в правое полушарие (и наоборот), имеет место у многих низших млекопитающих, таких как лошади и мыши, а у птиц и земноводных осуществляется даже полное перекрещивание зрительных нервов. У лошадей и мышей глаза расположены так, что они скорее направлены вбок, чем прямо вперед, в результате чего на большей части сетчатки правого глаза отображается правое зрительное поле, тогда как у приматов глаза направлены прямо вперед и на каждой сетчатке имеются отображения как правого, так и левого зрительного поля. Сделанное выше описание зрительных путей относится только к тем млекопитающим, у которых глаза смотрят более или менее прямо (как, например, у приматов) и поэтому видят почти один и тот же участок окружающего мира. Сходным образом устроена и слуховая система. Понятно, что каждое ухо способно слышать звуки, исходящие как из левой, так и из правой половины окружающего пространства. Подобно глазам, каждое ухо передает информацию о звуке примерно в равной степени в обе половины мозга. При этом в слуховой системе, как и в зрительной, процесс передачи информации латерализован: звук, доходящий до каждого уха от некоторого источника с правой стороны, анализируется в стволе мозга путем сравнения амплитуд и моментов прихода сигналов к тому и другому уху, и в результате реакция на этот звук формируется в основном в высших отделах левой половины мозга. Здесь мы говорим о начальных этапах переработки информации. Если справа от меня стоит человек, который словами или жестами побуждает меня сделать что-нибудь левой рукой, то сообщаемая им информация рано или поздно должна попасть в правое полушарие моего мозга. Однако первоначально сигналы должны прийти в слуховую или зрительную кору левого полушария, и только после этого они передаются в моторную кору правого полушария. Между прочим, никто не знает, почему правая половина окружающего пространства обычно проецируется в левое полушарие головного мозга. Из этого правила есть одно важное исключение — полушария мозжечка (отдел мозга, который в основном управляет движениями) получают входные сигналы главным образом от той же самой, а не противоположной стороны окружающего пространства. Это усложняет работу мозга, поскольку все волокна, соединяющие одно полушарие мозжечка с моторной корой в другом полушарии большого мозга, должны переходить с одной стороны мозга на другую. По поводу такой организации можно пока сказать лишь то, что она кажется загадочной.
Слоистая структура наружного коленчатого тела (НКТ)
Каждое из НКТ содержит по шесть клеточных слоев. Отдельный слой имеет толщину в несколько клеток (от 4 до 10 и более). Весь этот шестислойный сэндвич согнут таким образом, что его поперечный срез имеет вид, показанный на рис. 38. При переходе от сетчатки к НКТ простая схема, в которой каждый последующий слой клеток содержит проекцию предыдущего, становится более сложной. В НКТ проекции от сетчаток двух глаз объединяются, и два отдельных изображения, представленные на уровне ганглиозных клеток в сетчатках, проецируются на шесть слоев НКТ. Волокна от правого и левого глаза не конвергируют на одни и те же клетки НКТ — каждая из этих клеток получает сигналы только от какого-то одного глаза. Два множества клеток разнесены по отдельным слоям, так что в любом слое все клетки получают информацию только от одного глаза. Эти слои расположены таким образом, что проекции от правого и левого глаза чередуются. Так, в левом НКТ проекции располагаются в следующем порядке (от поверхности в глубину): левая, правая, левая, правая, правая, левая. Не совсем ясно, почему последовательность 5-го и 6-го слоев «перевернута» (иногда мне кажется, что это сделано для того, чтобы порядок проекций было труднее запомнить). У нас пока нет никакого вразумительного объяснения самого факта чередования проекций.
 Рис. 38. Левое наружное коленчатое тело макака. Ясно видны шесть клеточных слоев. Срез сделан параллельно фронтальной плоскости; он специально окрашен для выявления тел нейронов (каждое из них выглядит как точка).
Рис. 38. Левое наружное коленчатое тело макака. Ясно видны шесть клеточных слоев. Срез сделан параллельно фронтальной плоскости; он специально окрашен для выявления тел нейронов (каждое из них выглядит как точка).
В целом шестислойная нейронная структура имеет одну общую топографию для всех слоев. Левые половины обеих сетчаток проецируются в слои левого НКТ (рис. 39), а правые половины — в слои правого НКТ. Любая точка в одном слое НКТ соответствует некоторой точке в поле зрения того или другого глаза. Если двигаться вдоль слоя НКТ, то соответственная точка в поле зрения будет перемещаться по некоторой траектории, определяемой характером отображения зрительного поля на НКТ. Если же двигаться перпендикулярно слоям НКТ (например, вдоль черной штриховой линии на рис. 38), как двигался бы микроэлектрод, проходя через разные слои, то соответствующие рецептивные поля клеток оставались бы в одном и том же участке поля зрения; при этом наблюдалось бы только чередование проекций от разных глаз, за исключением, конечно, того места, где идут подряд две проекции от одного глаза. Таким образом, каждая половина поля зрения шестикратно отображается на каждое из НКТ, трижды для каждого глаза, а проекции в слоях НКТ располагаются в точности друг под другом.
 Рис. 39. При переходе от сетчатки к НКТ пространственная упорядоченность нейронов сохраняется, хотя на этом пути она временно исчезает, когда волокна собираются в пучок; в НКТ они снова «находят свои места».
Рис. 39. При переходе от сетчатки к НКТ пространственная упорядоченность нейронов сохраняется, хотя на этом пути она временно исчезает, когда волокна собираются в пучок; в НКТ они снова «находят свои места».
Наружное коленчатое тело выглядит как состоящее из двух частей. Его подразделяют на вентральные, или нижние, слои и четыре дорсальных, или верхних, слоя (вентральный — расположенный ближе к брюшной стороне тела, дорсальный — к спинной стороне). Вентральная часть НКТ образует особую структуру, так как клетки в соответствующих слоях отличаются от клеток в остальных четырех слоях — они крупнее и по-иному отвечают на зрительные стимулы. В то же время четыре слоя дорсальной части НКТ как гистологически, так и по своим электрофизиологическим свойствам сходны друг с другом. Поскольку величина клеток в этих двух отделах различна, вентральные слои стали называть крупноклеточными, а дорсальные — мелкоклеточными. Волокна, выходящие из шести слоев НКТ, объединяются в один широкий пучок, называемый зрительной радиацией, который идет вверх до первичной зрительной коры (см. рис. 35). Здесь эти волокна равномерно расходятся и перераспределяются так, что образуется целостная проекция с топографической организацией (это аналогично распределению волокон зрительного нерва при входе его в НКТ). И вот, наконец, мы подходим к коре.
Ответы клеток в коре
Главная тема этой главы — вопрос о том, как клетки в первичной зрительной коре отвечают на зрительные стимулы. Рецептивные поля нейронов НКТ имеют такую же организацию (разделение на центр и периферию), как и рецептивные поля ганглиозных клеток сетчатки, которые посылают свои аксоны к клеткам НКТ. Подобно ганглиозным клеткам сетчатки, нейроны НКТ различаются между собой главным образом свойствами рецептивного поля (on- или off-центр, местоположение в поле зрения) и особенностями ответов на цветовые стимулы. Возникает вопрос: а как обстоит дело с корковыми нейронами? Сходны ли они с клетками НКТ, посылающими в кору свои аксоны, или же у них появляются какие-то новые особенности? Ответ, как читатель уже должен догадаться, такой: корковые клетки действительно обладают новыми качествами, причем настолько необычными, что вплоть до 1958 года, когда их впервые стали изучать с помощью сложных световых стимулов, никто не мог даже приблизительно предсказать эти свойства. Первичная зрительная кора (стриарная кора) представляет собой слой клеток толщиной 2 мм и площадью в несколько квадратных дюймов.[1] Для того чтобы дать представление о размерах этой нейронной структуры, можно привести такие цифры: если НКТ содержит полтора миллиона клеток, то стриарная кора — около 200 миллионов клеток. Анатомическая структура стриарной коры удивительно сложна, однако нет необходимости знать ее детали, чтобы понять, каким образом преобразуется здесь поступающая зрительная информация. Более подробно строение этого отдела будет рассмотрено в следующей главе, где будет обсуждаться вопрос о его функциональной архитектуре. Как я уже говорил, процесс переработки информации в коре состоит из нескольких этапов. На первом этапе большинство клеток дает такие же ответы, как клетки НКТ. Рецептивные поля этих клеток обладают круговой симметрией. Это означает, что линия или граница (перепад освещенности) вызывает один и тот же ответ вне зависимости от ее ориентации. Регистрировать электрическую активность корковых клеток этого уровня непросто, так как они очень малы и расположены близко друг к другу. Пока еще не ясно, отличаются ли вообще ответы этих корковых клеток от ответов клеток НКТ (точно так же как не ясно, отличаются ли реакции клеток НКТ от ответов ганглиозных клеток сетчатки). Сложность гистологического строения НКТ и коры позволяет думать, что между ними должны быть какие-то различия и что их можно будет выявить, если знать, в чем их следует искать; однако узнать это может оказаться трудным делом. Положение еще больше усложняется, когда мы переходим к ответам клеток на следующем корковом уровне. Вероятно, эти клетки должны получать входные сигналы от нейронов предыдущего уровня, обладающих рецептивными полями с центром и периферией. Вначале нам было совсем не просто выяснить, на какие зрительные стимулы отвечают эти клетки второго коркового уровня. В то время (конец 50-х годов) лишь очень немногие исследователи пытались регистрировать ответы одиночных нейронов зрительной коры. Те, кто это делал, получали противоречивые результаты. Они нашли, что клетки в зрительной коре работают, видимо, почти так же, как и в сетчатке, — были найдены как on-, так и off-клетки. Кроме того, был обнаружен еще один класс клеток, которые, казалось, вообще не отвечали на световые стимулы. Простота выявленных физиологических свойств корковых клеток на фоне просто дьявольской сложности морфологии коры ставила исследователей в тупик.
 Рис. 40. Окрашенный по Гольджи срез первичной зрительной коры, на котором видно больше дюжины пирамидных клеток (но это лишь очень малая доля нейронов, содержащихся в срезе). Величина изображенного участка по вертикали — около 1 мм. (Темная полоса у правого края — кровеносный сосуд.)
Рис. 40. Окрашенный по Гольджи срез первичной зрительной коры, на котором видно больше дюжины пирамидных клеток (но это лишь очень малая доля нейронов, содержащихся в срезе). Величина изображенного участка по вертикали — около 1 мм. (Темная полоса у правого края — кровеносный сосуд.)
Сегодня это очень просто объяснить. Во-первых, применялись неадекватные стимулы — для активации клеток коры по привычке просто освещали всю сетчатку рассеянным светом, хотя Куффлер еще за десять лет до того показал, что такая стимуляция даже для нейронов сетчатки далеко не оптимальна. Для большинства же корковых нейроновдиффузный засвет сетчатки — не только не оптимальный, но вообще полностью неэффективный стимул. Если многие клетки наружных коленчатых тел еще реагируют, хотя и слабо, на диффузный засвет, то корковые клетки, даже относящиеся к первому корковому уровню и сходные с клетками НКТ, практически не отвечают на такую стимуляцию. Таким образом, первая приходящая в голову мысль, что для активации зрительных нейронов лучше всего стимулировать все рецепторы сетчатки, оказалась совершенно неверной. Во-вторых (и это еще парадоксальнее), оказалось, что те корковые клетки, которые давали on- или off-ответы, на самом деле были не клетками, а просто аксонами клеток НКТ. Настоящие корковые нейроны в этом случае вообще не отвечали на стимуляцию! Они считали ниже своего достоинства обращать внимание на такой примитивный стимул, как диффузный свет. Именно так обстояло дело в 1958 году, когда Торстену Визелу и мне удалось осуществить одно из первых успешных отведений электрической активности коры у кошки. В этих опытах положение кончика микроэлектрода в корковой ткани было настолько стабильным, что можно было прослушивать активность одной и той же клетки в течение примерно девяти часов. Чтобы вызывать разряды клетки, мы использовали все возможные и невозможные средства — разве что сами не вставали на голову. (Как большинство корковых клеток, время от времени найденная клетка давала спонтанный разряд, но мы пытались убедить друг друга, что разряд был вызван именно нашими стимулами, и в результате потратили впустую много времени.) После нескольких часов безуспешных попыток у нас возникло смутное впечатление, что подача света на одну определенную область сетчатки вызывала некоторую реакцию, и мы сосредоточили свои усилия именно на этой области. В качестве стимулов мы в основном использовали круглые белые и черные пятнышки. Чтобы создать черное пятно, мы обычно брали предметное стекло размером 2,5×5 см, на которое был наклеен непрозрачный кружок; стекло вставлялось в специальное проекционное устройство, которое Сэмюэл Талбот сконструировал для подачи изображений на сетчатку. Для предъявления белых пятен мы брали медную пластинку такого же размера с небольшим отверстием (в те времена затраты на проведение исследований были гораздо меньшими). По прошествии примерно пяти часов напряженной работы нам неожиданно показалось, что, когда время от времени клетка случайно давала реакцию, эта реакция скорее была связана не с черным пятнышком, нанесенным на стекло, а с самим этим стеклом. В конце концов нам удалось установить, что реакцию клетки вызывала слабая, но четкая тень от края стеклянной пластинки, когда ее задвигали в окошко проекционного устройства. Вскоре мы убедились, что этот край работал как стимул только тогда, когда тень от него пересекала определенный участок сетчатки, причем эта тень должна была иметь вполне определенную ориентацию. Самым удивительным было резкое различие результатов в двух случаях — когда ориентация стимула была оптимальной (тогда разряд напоминал пулеметную очередь) и когда мы изменяли ориентацию стимула или производили тотальную засветку глаза яркой вспышкой (в этом случае никакой реакции не было).
 Рис. 41. Ответы одной из первых клеток, чувствительных к ориентации, которые Торстен Визел и я обнаружили в стриарной коре кошки в 1958 году. Данная клетка отвечала исключительно на стимул в виде движущейся щели с ориентацией, соответствующей положению часовой стрелки в 11 часов; при этом она реагировала на движение стимула вправо вверх и не реагировала на движение влево вниз.
Рис. 41. Ответы одной из первых клеток, чувствительных к ориентации, которые Торстен Визел и я обнаружили в стриарной коре кошки в 1958 году. Данная клетка отвечала исключительно на стимул в виде движущейся щели с ориентацией, соответствующей положению часовой стрелки в 11 часов; при этом она реагировала на движение стимула вправо вверх и не реагировала на движение влево вниз.
Этот феномен был обнаружен самым первым и направил нас по ложному пути. Дело в том, что, как нарочно, обнаруженный тип клеток был как раз тот, который мы впоследствии назвали классом сложных клеток, а они относятся к уровню на две ступени выше, чем первый корковый уровень клеток с рецептивными полями, разделенными на центр и периферию. Хотя сложные клетки — наиболее распространенный тип клеток в стриарной коре, в их свойствах трудно разобраться, не познакомившись сначала с клетками предыдущего, промежуточного уровня. Действительно, в отличие от клеток первого уровня, имеющих рецептивные поля с центром и периферией, у обезьяны клетки более высоких уровней проявляют совершенно иные реакции. Обычно точечные световые стимулы вызывают лишь слабую реакцию этих клеток или вообще неэффективны. Для того чтобы вызвать ответ клетки, сначала нужно найти надлежащий участок зрительного поля для предъявления стимула, т.е. найти соответствующий участок экрана, перед которым находится животное. Иными словами, сначала мы должны выявить рецептивное поле данной клетки. После этого выясняется, что наиболее эффективным стимулом для данной клетки служит линия, движущаяся в рецептивном поле в направлении, перпендикулярном ориентации линии. Такой линией могла быть узкая полоса света на темном фоне (щель), темная полоса на светлом фоне или же прямолинейная граница между темной и светлой областями. Некоторые клетки отдавали предпочтение, часто очень сильно выраженное, какому-то одному из этих трех стимулов, другие отвечали с примерно одинаковой интенсивностью на стимулы всех трех типов. Решающее значение имела ориентация линии — чаще всего клетка лучше всего реагировала на определенную оптимальную ориентацию, причем интенсивность ответа (число импульсов, возникавших при пересечении стимулом рецептивного поля) заметно снижалась при отклонении ориентации в любую сторону от оптимальной на 10–20 градусов; при еще большем отклонении реакция круто снижалась до нулевого уровня (см. рис. 41). Указанная цифра 10–20 градусов может показаться очень приблизительной. Вспомните, однако, что разница в положении часовой стрелки, показывающей «час» и «два», еще больше — 30 градусов. Когда ориентация стимула отличается от оптимальной на 90 градусов, типичная клетка, избирательно чувствительная к ориентации, перестает отвечать вообще. В отличие от клеток на более низких уровнях зрительной системы нейроны, избирательно чувствительные к ориентации стимула, гораздо лучше отвечают на движущиеся, чем на неподвижные линии. Именно поэтому (см. рис. 41) при стимуляции таких нейронов мы использовали линии, движущиеся через рецептивное поле. Если использовать в качестве стимула неподвижную мелькающую (периодически вспыхивающую) линию, то зачастую клетка дает слабый ответ, и в этом случае предпочтительной оказывается такая же ориентация, как и при движущейся линии. Многие клетки (вероятно, треть всей популяции) дают еще один характерный вид ответа на движущийся стимул. Вместо того чтобы давать один и тот же импульсный разряд независимо от направления движения, такие клетки отвечают более энергично при одном определенном направлении. Бывает даже так, что движение в одну сторону вызывает сильно выраженный ответ, а при движении в противоположную сторону нет вообще никакой реакции (это показано на рис. 41). За один эксперимент можно оценить реакции 200–300 клеток, если после полного исследования одной клетки просто продвигать микроэлектрод дальше, до следующей клетки. Недостаток такой методики в том, что в одном прогоне можно исследовать только клетки, лежащие в коре на одной прямой линии: как только вы ввели тончайший микроэлектрод в корковую ткань, вы уже не можете сдвинуть его в поперечном направлении, не повредив при этом сам электрод или еще более нежную нервную ткань. С помощью такой методики регистрации самое большее, что мы можем сделать — это исследовать при одной проходке микроэлектрода примерно 50 клеток на миллиметр. Когда мы исследуем ориентационную избирательность нескольких сотен или тысяч клеток, оказывается, что все ориентации стимула встречаются примерно одинаково часто — вертикальная, горизонтальная и все промежуточные, наклонные ориентации. Если учесть характерные особенности окружающего нас мира, в который входят как деревья, так и линия горизонта, то возникнет вопрос: нет ли каких-нибудь выделенных ориентаций, таких как вертикаль и горизонталь, которые встречались бы чаще, чем другие? При попытках ответить на этот вопрос в разных лабораториях получали несколько разные результаты. Однако все исследователи соглашаются, что, если такие предпочтения действительно имеют место, они должны быть очень малы — настолько малы, что для выявления их требуется статистическая обработка данных. А в этом случае они вряд ли имеют какое-либо значение! В стриарной коре обезьян примерно 70–80% клеток обладают свойствами ориентационной избирательности. Что касается кошек, то у них, по-видимому, все корковые клетки чувствительны к ориентации стимула, даже те, которые имеют прямые входы от наружных коленчатых тел. Мы обнаружили заметные различия среди ориентационно-специфичных клеток, причем не столько в оптимальной ориентации стимула или в положении рецептивного поля на сетчатке, сколько в характере поведения клеток. Наиболее существенно различие между двумя классами нейронов — простыми и сложными клетками. Как можно догадаться по их названиям, клетки этих двух классов различаются по сложности своих ответных реакций. Поэтому мы сделали естественное предположение о том, что клетки с более простым поведением расположены в нейронной структуре коры ближе к ее входу.
Простые клетки
В большинстве случаев по ответам простых клеток на стимул в виде маленького светового пятна можно предсказать их реакцию на стимул сложной формы. Каждая из простых клеток, подобно ганглиозным клеткам сетчатки, клеткам НКТ и корковым клеткам с центрально-симметричными рецептивными полями, имеет небольшое четко очерченное рецептивное поле. Предъявление в пределах этого рецептивного поля стимула в виде светового пятнышка вызывает либо on-, либо off-реакцию в зависимости от того, в какой именно участок рецептивного поля подан стимул. Различие между простыми клетками и клетками предыдущих уровней заключается в конфигурации зон возбуждения и торможения. На предыдущих уровнях это центрально-симметричная конфигурация — имеется одна центральная on- или off-зона (возбуждающая или тормозная) и окружающая ее со всех сторон кольцевая зона с противоположными свойствами (тормозная или возбуждающая). Простые клетки коры более сложны. Зоны возбуждения и торможения в их рецептивных полях всегда разделены одной прямой линией или двумя параллельными линиями (рис. 42). Чаще всего встречается такая конфигурация, когда к длинной и узкой возбуждающей зоне с двух сторон примыкают более широкие тормозные зоны (рис. 42, А).
 Рис. 42. Карты трех типичных рецептивных полей простых клеток. Оптимальными стимулами служили: для клетки А — светлая полоса против возбуждающей области (+); для клетки Б — темная линия, покрывающая тормозную зону (–); для клетки В — резкая граница «темное — светлое», совпадающая с границей между возбуждающей и тормозной зонами.
Рис. 42. Карты трех типичных рецептивных полей простых клеток. Оптимальными стимулами служили: для клетки А — светлая полоса против возбуждающей области (+); для клетки Б — темная линия, покрывающая тормозную зону (–); для клетки В — резкая граница «темное — светлое», совпадающая с границей между возбуждающей и тормозной зонами.
Для того чтобы проверить предполагаемую карту рецептивного поля, составленную путем тестирования маленьким световым пятном, мы попробовали использовать в качестве стимулов другие конфигурации. Вскоре мы выяснили, что чем большую долю той или иной зоны рецептивного поля покрывает данный стимул, тем сильнее выражено возбуждение клетки или ее торможение. Иными словами, имеет место пространственная суммация локальных воздействий. Мы обнаружили также явление антагонизма — взаимного погашения локальных воздействий при одновременной стимуляции возбуждающей и тормозной зоны. Таким образом, для клетки с рецептивным полем, изображенным на рис. 42, А, наиболее подходящим будет стимул в виде узкой полосы, расположенной в рецептивном поле так, чтобы она точно совпадала с возбуждающей зоной и не заходила в тормозную зону (см. рис. 43). Даже незначительное изменение ориентации этой полосы приведет к уменьшению эффективно действующей площади зоны возбуждения и затронет также тормозную зону; в результате частота разряда в ответе клетки уменьшится.
 Рис. 43. Стимулы разных конфигураций вызывают различные реакции клетки с рецептивным полем такого типа, как А на рис. 42. Отрезком жирной линии внизу указан период (1 секунда), когда был включен стимул — светлая полоса. В первом случае (верхняя запись) показан ответ клетки на полосу оптимальных размеров, положения и ориентации. Во втором случае та же самая полоса покрывает только часть тормозной зоны (поскольку эта клетка не обладает спонтанной активностью, которая могла бы подавляться при торможении, здесь виден только разряд клетки при выключении стимула). В третьем случае полоса ориентирована так, что покрывает только малую часть возбуждающей зоны и соответственно малую часть тормозной зоны, и поэтому клетка не отвечает вообще. На нижней записи показан случай равномерного освещения всего рецептивного поля: ответа здесь тоже нет.
Рис. 43. Стимулы разных конфигураций вызывают различные реакции клетки с рецептивным полем такого типа, как А на рис. 42. Отрезком жирной линии внизу указан период (1 секунда), когда был включен стимул — светлая полоса. В первом случае (верхняя запись) показан ответ клетки на полосу оптимальных размеров, положения и ориентации. Во втором случае та же самая полоса покрывает только часть тормозной зоны (поскольку эта клетка не обладает спонтанной активностью, которая могла бы подавляться при торможении, здесь виден только разряд клетки при выключении стимула). В третьем случае полоса ориентирована так, что покрывает только малую часть возбуждающей зоны и соответственно малую часть тормозной зоны, и поэтому клетка не отвечает вообще. На нижней записи показан случай равномерного освещения всего рецептивного поля: ответа здесь тоже нет.
На рис. 42, Б и В показаны рецептивные поля простых клеток двух других типов, которые отвечают наилучшим образом на темные линии и на прямолинейные границы светлого и темного; при этом чувствительность клеток к ориентации стимула остается примерно такой же, как у клеток первого типа. Клетки всех трех типов совсем не реагируют на стимул в виде диффузного освещения. Такое взаимное погашение процессов возбуждения и торможения напоминает реакцию нейтрализации кислоты основанием, которую выполняют студенты в лабораторных работах по химии. Таким образом, уже на этом корковом уровне отмечается большое многообразие нейронов. Если взять класс простых клеток, то у них встречаются три или четыре разных типа рецептивных полей, причем имеются клетки, настроенные на любую из возможных ориентаций стимула, и клетки с рецептивным полем в любом участке поля зрения. Величина рецептивных полей простых клеток зависит от их расстояния от центральной ямки. Однако в одной и той же зоне сетчатки тоже есть некоторые различия в размерах рецептивных полей. Самые маленькие рецептивные поля, расположенные в центральной ямке и около нее, имеют величину примерно 0,25×0,25°. Что касается клеток типа, показанного на рис. 42, А и Б, то ширина центральной зоны составляет у них не более нескольких угловых минут. Эта величина совпадает с минимальным диаметром рецептивных полей ганглиозных клеток сетчатки или клеток НКТ. Если же взять область далекой периферии, то размеры рецептивных полей простых клеток здесь могут достигать 1×1°.
 Рис. 44. Возможная схема связей, определяющих рецептивное поле простой клетки. Четыре клетки образуют возбуждающие синаптические связи с клеткой более высокого порядка. Каждая из клеток низшего порядка имеет рецептивное поле с радиальной симметрией, возбуждающим центром и тормозной периферией (это показано на схеме слева). Центры этих рецептивных полей лежат вдоль прямой линии. Если мы предположим, что с данной простой клеткой соединено много клеток, имеющих рецептивные поля с центром и периферией, причем центры этих рецептивных полей перекрываются и лежат на одной прямой, то рецептивное поле простой клетки будет состоять из длинной, узкой возбуждающей зоны и тормозных флангов. Избегая специальной терминологии, можно просто сказать, что небольшое светлое пятно в любом месте этого длинного и узкого прямоугольника будет приводить к сильному возбуждению одной или нескольких клеток с круглыми рецептивными полями и в результате — активировать, хотя и слабо, простую клетку. Если же стимулом будет длинная узкая полоса, способная активировать все клетки с круглыми рецептивными полями, то это приведет к сильной реакции простой клетки
Рис. 44. Возможная схема связей, определяющих рецептивное поле простой клетки. Четыре клетки образуют возбуждающие синаптические связи с клеткой более высокого порядка. Каждая из клеток низшего порядка имеет рецептивное поле с радиальной симметрией, возбуждающим центром и тормозной периферией (это показано на схеме слева). Центры этих рецептивных полей лежат вдоль прямой линии. Если мы предположим, что с данной простой клеткой соединено много клеток, имеющих рецептивные поля с центром и периферией, причем центры этих рецептивных полей перекрываются и лежат на одной прямой, то рецептивное поле простой клетки будет состоять из длинной, узкой возбуждающей зоны и тормозных флангов. Избегая специальной терминологии, можно просто сказать, что небольшое светлое пятно в любом месте этого длинного и узкого прямоугольника будет приводить к сильному возбуждению одной или нескольких клеток с круглыми рецептивными полями и в результате — активировать, хотя и слабо, простую клетку. Если же стимулом будет длинная узкая полоса, способная активировать все клетки с круглыми рецептивными полями, то это приведет к сильной реакции простой клетки
Даже теперь, спустя двадцать лет, мы все еще не знаем, как устроены входные для корковых клеток цепи, от которых зависят специфические реакции этих клеток. Предложен ряд правдоподобных схем, и вполне может оказаться, что одна из этих схем или какая-то их комбинация окажется верной. Свойства простых клеток могут определяться нейронами предшествующего уровня с круглыми рецептивными полями; проще всего предположить, что простые клетки имеют прямые возбуждающие входы от многих клеток предыдущего уровня — таких, у которых центры рецептивных полей лежат в зрительном поле на одной прямой линии (рис. 44). Несколько труднее предложить гипотетическую схему для клеток, избирательно реагирующих на границы темного и светлого (см. рис. 42, В). Возможен такой вариант: данная простая клетка имеет входы от двух наборов клеток предыдущего уровня, у которых центры рецептивных полей расположены с двух сторон от одной линии — по одну сторону клетки с on-центрами, а по другую с off-центрами, причем все эти входы возбуждающие. Во всех таких гипотетических схемах возбуждающий вход от клетки с on-центром по логике вещей эквивалентен тормозному входу от клетки с off-центром, при условии, что клетка с off-центром обладает спонтанной активностью. Выяснение действительного механизма, определяющего реакции простых клеток, — задача непростая. Относительно данной простой клетки нужно будет узнать, от какого рода клеток она получает входные сигналы, например выяснить для каждой из предшествующих клеток структуру рецептивного поля, его местоположение и ориентацию (если она имеется), тип центра (on- или off-), а также характер посылаемых сигналов — возбуждающие они или тормозные. Поскольку методов получения таких сведений пока нет, мы вынуждены пользоваться косвенными подходами, а это увеличивает вероятность ошибки. Схема, представленная на рис. 44, кажется мне наиболее правдоподобной, так как она самая простая.
Сложные клетки
Сложные клетки соответствуют следующему уровню (или уровням) зрительного анализа. Они наиболее многочисленны в стриарной коре и составляют здесь, вероятно, около трех четвертей всей популяции нейронов. Первая из клеток, чувствительных к ориентации стимула, исследованная Визелом и мною, — та, которая отвечала на стимул в виде края стеклянного слайда, — почти наверное была сложной клеткой. Общим свойством сложных и простых клеток является способность реагировать только на линии, ориентированные определенным образом. Сложные клетки, так же как и простые, отвечают на стимулы, предъявляемые в ограниченном участке поля зрения. От простых они отличаются тем, что реакции их нельзя объяснить формой и распределением возбуждающих и тормозных зон в рецептивном поле. Включение или выключение небольшого неподвижного пятна в пределах рецептивного поля редко вызывает ответ клетки. Даже на надлежащим образом ориентированную неподвижную полосу или границу клетка чаще всего не реагирует или дает лишь слабый, быстро затухающий ответ одного и того же типа как при включении, так и при выключении стимула. Однако, если должным образом ориентированная линия перемещается через рецептивное поле, возникает хорошо выраженный длительный разряд импульсов. Этот разряд начинается в момент, когда линия входит в рецептивное поле, и продолжается до тех пор, пока она не выйдет за его пределы (см. рис. 41, где показана схема ответа). Напротив, для того, чтобы вызвать длительный разряд простой клетки, нужно предъявить надлежащим образом ориентированную неподвижную линию в определенном участке рецептивного поля. Если же использовать движущуюся линию, то возникает только кратковременная реакция в тот момент, когда линия пересекает границу тормозной и возбуждающей зон, или же в то время, когда линия проходит через возбуждающую зону рецептивного поля. Те сложные клетки, которые способны реагировать на неподвижные световые «щели», полоски или границы, дают импульсный разряд независимо от того, в каком месте рецептивного поля расположен стимул, лишь бы ориентация его была подходящей. Однако те же стимулы совершенно неэффективны, если их ориентация далека от оптимальной (рис. 46).
 Рис. 45. Активность этой клетки из слоя 5 стриарной коры кошки была записана в 1973 году с помощью внутриклеточного электрода Д. Эссеном и Дж. Келли из Гарвардской медицинской школы. Было проведено также картирование ее сложного рецептивного поля. Затем путем инъекции красителя проционового желтого выяснили, что это была пирамидная клетка.
Рис. 45. Активность этой клетки из слоя 5 стриарной коры кошки была записана в 1973 году с помощью внутриклеточного электрода Д. Эссеном и Дж. Келли из Гарвардской медицинской школы. Было проведено также картирование ее сложного рецептивного поля. Затем путем инъекции красителя проционового желтого выяснили, что это была пирамидная клетка.
Схемы на рис. 46 (для сложной клетки) и на рис. 43 (для простой клетки) демонстрируют существенное различие между двумя типами клеток: у простой клетки реакцию вызывает оптимально ориентированная линия лишь в очень узком диапазоне положений, а у сложной такая линия вызывает ответ, в каком бы участке рецептивного поля она ни предъявлялась. Это различие связано с существованием четко очерченных возбуждающих и тормозных зон в рецептивном поле простой клетки и с отсутствием их в рецептивном поле сложной клетки. Сложная клетка дает пример генерализации (нелокальности) ответа на линию в пределах более обширной области. В целом у сложных клеток рецептивные поля несколько больше, чем у простых, но не намного. У макака в области центральной ямки чаще всего встречаются рецептивные поля сложных клеток величиной примерно 0,5×0,5°. В этой области сетчатки оптимальные размеры стимула как для простых, так и для сложных клеток составляют около двух угловых минут. Таким образом, «разрешающая способность» сложных клеток такая же, как у простых клеток. Как и в отношении простых клеток, мы как следует не знаем, как организована система связей, передающая сигналы сложным клеткам. Однако и здесь нетрудно предложить несколько возможных схем их организации. Согласно простейшей из таких схем, сложная клетка получает входные сигналы от множества простых клеток, рецептивные поля которых имеют одну и ту же ориентацию, но размещены, частично перекрывая друг друга, по всему полю сложной клетки, как показано на рис. 47. Если связи от простых клеток к сложным возбуждающие, то всякий раз, когда в рецептивное поле сложной клетки попадает стимул в виде линии, возбуждаются некоторые простые клетки. В результате будет возбуждаться и сложная клетка.
 Рис. 46. Длинная и узкая полоса света вызывает реакцию сложной клетки независимо от того, в каком месте рецептивного поля она предъявлена, если только ее ориентация оптимальна (три верхние записи). Если ориентация полосы отличается от оптимальной, клетка реагирует слабее или не отвечает вовсе (нижняя запись).
Рис. 46. Длинная и узкая полоса света вызывает реакцию сложной клетки независимо от того, в каком месте рецептивного поля она предъявлена, если только ее ориентация оптимальна (три верхние записи). Если ориентация полосы отличается от оптимальной, клетка реагирует слабее или не отвечает вовсе (нижняя запись).
 Рис. 47. Схема связей, которая позволила бы объяснить наблюдаемые свойства сложной клетки. Мы предполагаем, что (как и на рис. 44) к одной сложной клетке могут приходить возбуждающие сигналы от большого числа простых клеток (здесь показаны только три). Каждая простая клетка наилучшим образом отвечает на вертикальную границу между светлым (слева) и темным (справа) участками. Предполагается, что рецептивные поля простых клеток разбросаны в пределах прямоугольника и перекрываются. Если стимул в виде такой границы подается в любое место прямоугольника, то некоторое число простых клеток активируется и это в свою очередь вызывает ответ сложной клетки. Из-за эффекта адаптации синапсов только движущийся стимул будет вызывать непрерывное возбуждение сложной клетки.
Рис. 47. Схема связей, которая позволила бы объяснить наблюдаемые свойства сложной клетки. Мы предполагаем, что (как и на рис. 44) к одной сложной клетке могут приходить возбуждающие сигналы от большого числа простых клеток (здесь показаны только три). Каждая простая клетка наилучшим образом отвечает на вертикальную границу между светлым (слева) и темным (справа) участками. Предполагается, что рецептивные поля простых клеток разбросаны в пределах прямоугольника и перекрываются. Если стимул в виде такой границы подается в любое место прямоугольника, то некоторое число простых клеток активируется и это в свою очередь вызывает ответ сложной клетки. Из-за эффекта адаптации синапсов только движущийся стимул будет вызывать непрерывное возбуждение сложной клетки.
Обычно в ответ на неподвижную линию сложная клетка дает короткий импульсный разряд (даже если стимул остается включенным). В этом случае мы говорим, что происходит адаптация ответа. Если же перемещать линию в рецептивном поле сложной клетки, наблюдается непрерывный разряд: адаптация преодолевается в результате последовательного срабатывания новых простых клеток. Читатель, должно быть, заметил, что обе приведенные схемы связей — от клеток с круглыми рецептивными полями к простым клеткам (рис. 44) и от простых клеток к сложным (рис. 47) — предполагают использование возбуждающих связей. Однако в этих двух случаях процессы возбуждения должны быть совершенно различными. В первой из этих схем требуется суммирование одновременных сигналов от клеток с круглыми рецептивными полями, лежащих на одной линии. Во второй же схеме для активации сложной клетки движущимся стимулом необходимо последовательное возбуждение множества простых клеток. Было бы интересно выяснить, с какими морфологическими различиями (если они есть) связано это различие в механизме суммации.
Дирекциональная избирательность
Многие сложные клетки лучше реагируют на движение стимула в одном направлении, чем в противоположном. Различие в реакции часто бывает весьма резким — при одном направлении движения возникает энергичный ответ, а при обратном направлении клетка вообще не отвечает (рис. 48). Как выяснилось, примерно 10–20% клеток в верхних слоях стриарной коры проявляют заметную дирекциональную избирательность. Остальные клетки, по-видимому, не обладают такой избирательностью — мы тщательно исследовали ответы клеток с помощью ЭВМ, пытаясь обнаружить хотя бы небольшое различие в ответах на движение стимула в противоположных направлениях. Таким образом, существуют, видимо, два разных класса клеток — один из них четко проявляет дирекциональную избирательность, другому она не свойственна.
 Рис. 48. Реакции данной сложной клетки на движение оптимально ориентированной полосы в противоположных направлениях различны. Длительность каждой записи около 2 секунд. (Для таких клеток не особенно важно, насколько быстро движется полоса; обычно клетка не отвечает только при очень быстром движении, когда полоса кажется размытой, или же при таком медленном, что его вообще трудно заметить.)
Рис. 48. Реакции данной сложной клетки на движение оптимально ориентированной полосы в противоположных направлениях различны. Длительность каждой записи около 2 секунд. (Для таких клеток не особенно важно, насколько быстро движется полоса; обычно клетка не отвечает только при очень быстром движении, когда полоса кажется размытой, или же при таком медленном, что его вообще трудно заметить.)
Если прослушивать импульсную реакцию клетки с сильно выраженной дирекциональной избирательностью, то создается впечатление, что при движении линии в одном направлении стимул как бы резко подталкивает клетку и заставляет ее разряжаться, а при движении в противоположном направлении происходит как бы сбой и стимул становится неэффективным (это напоминает ситуацию, когда вы, заводя часы, поворачиваете головку заводного механизма назад и раздается характерный треск). Мы не знаем, как устроены входные сети таких клеток с дирекциональной избирательностью. Возможно, что на вход такой клетки подключены простые клетки, реакции которых на движение стимула в противоположных направлениях неравнозначны, асимметричны. Рецептивные поля таких простых клеток асимметричны, как, например, поле, представленное на рис. 42, В. Другую возможную схему предложили в 1965 году Г. Барлоу и У. Левик для объяснения дирекциональной избирательности некоторых клеток в сетчатке кролика — клеток, которых, по-видимому, нет у обезьяны. Если применить их схему к сложным клеткам коры, то нужно предположить наличие промежуточного слоя клеток, расположенных между простыми и сложными клетками (как на рис. 49). Представим себе, что клетка из промежуточного слоя имеет возбуждающий вход от одной простой клетки и тормозный вход — от другой клетки, рецептивное поле которой непосредственно примыкает к предыдущему, и притом всегда с одной стороны. Предположим далее, что тормозный путь включает некоторую задержку, возможно, за счет включения еще одной промежуточной клетки. В таком случае, если стимул движется в одном направлении, скажем справа налево (как на рис. 49, иллюстрирующем модель Барлоу и Левика), то промежуточная клетка возбуждается сигналом с одного из своих входов как раз в тот момент, когда действует торможение, приходящее от другой клетки, рецептивное поле которой только что пересек стимул. Возбуждающий и тормозный эффекты нейтрализуют друг друга, и в результате клетка не дает импульсного разряда. Если же стимул движется в обратном направлении, торможение возникает слишком поздно для того, чтобы предотвратить импульсный разряд. Если множество таких промежуточных клеток конвергирует на какую-то клетку, расположенную на следующем, третьем, уровне, то эта клетка будет обладать свойствами сложной клетки с дирекциональной избирательностью.
 Рис. 49. Эту схему предложили X. Барлоу и У. Левик для объяснения свойства дирекциональной чувствительности. Синапсы, которые красные клетки образуют на зеленых клетках, — возбуждающие, а синапсы, образуемые на белых клетках, — тормозные. Мы предполагаем, что три белые клетки (внизу) конвергируют на одну «главную» клетку.
Рис. 49. Эту схему предложили X. Барлоу и У. Левик для объяснения свойства дирекциональной чувствительности. Синапсы, которые красные клетки образуют на зеленых клетках, — возбуждающие, а синапсы, образуемые на белых клетках, — тормозные. Мы предполагаем, что три белые клетки (внизу) конвергируют на одну «главную» клетку.
У нас нет прямых доказательств правильности тех или иных иерархических схем, объясняющих наблюдаемые реакции клеток, т.е. схем, в которых клетки на каждом следующем уровне организуются на основе элементов предыдущего уровня. Тем не менее у нас есть веские основания считать, что нервная система организована по иерархическому принципу. Одно из самых серьезных оснований для такого заключения связано с морфологией — например, у кошек простые клетки сосредоточены в четвертом слое стриарной коры, т.е. в том слое, который получает входы от клеток наружных коленчатых тел, тогда как сложные клетки располагаются в слоях коры выше и ниже — на одно или два синаптических переключения дальше от входа. Таким образом, хотя мы не можем дать точную схему связей на каждом уровне, у нас есть достаточные основания полагать, что такая схема существует. Главная причина того, что в сложных клетках мы видим результат определенной организации клеток с круглыми рецептивными полями с центром и периферией, — это очевидная необходимость производить переработку информации логически в два этапа. Я хотел бы подчеркнуть слово логически, поскольку в целом нужное преобразование, по-видимому, могло бы быть физически осуществлено и в один этап — если суммировать на отдельных разветвлениях дендритов сложных клеток входные сигналы от клеток с круглыми рецептивными полями. В этом случае каждое разветвление выполняло бы функции отдельной простой клетки, посылая к телу клетки (и соответственно к аксону) сигналы электротоническим путем (с помощью пассивного электрического процесса) всякий раз, когда какая-то линия попадет в определенную зону соответствующего рецептивного поля. В этом случае такая клетка сама проявляла бы свойства сложной клетки. Однако само наличие простых клеток заставляет думать, что мы не должны строить таких сложных воображаемых конструкций.
Значение клеток, чувствительных к движению (включая некоторые комментарии к тому, как мы «видим»)
Почему столь многочисленны клетки, чувствительные к движению? Прежде всего приходит в голову, что такие клетки сигнализируют о том, имеется ли в поле зрения движущийся объект. Для животных, в том числе и для нас с вами, изменения внешнего мира гораздо важнее, чем подтверждения его неизменности, идет ли речь о выживании хищника или жертвы. Поэтому не удивительно, что большинство корковых клеток лучше реагирует на движущиеся объекты. Следуя этой логике, вы должны теперь усомниться — как же тогда мы проводим анализ неподвижных зрительных сцен, если в интересах чувствительности к движению так много клеток, настроенных на определенную ориентацию стимула, нечувствительно к неподвижным контурам. Чтобы получить ответ на этот вопрос, нужно сделать небольшое отступление и остановиться на некоторых важных, но противоречащих интуиции данных относительно того, как мы «видим». Прежде всего естественно было бы ожидать, что при обследовании окружающего мира наши глаза будут плавно сканировать всю сцену непрерывными движениями. На самом же деле при фиксации какого-то объекта происходит вот что: сначала мы устанавливаем глаза так, чтобы изображение этого объекта попало в область центральной ямки обоих глаз, затем мы удерживаем глаза в таком положении в течение короткого времени, скажем полсекунды, потом глаза скачком перемещаются в новую позицию и фиксируют новую мишень, которая находится где-то в другом месте зрительного поля и привлекает к себе внимание тем, что несколько сдвигается относительно фона или имеет какую-то интересную форму. Во время такого скачка, или саккады, скорость движения глаз столь велика, что зрительная система не успевает отреагировать на перемещение изображения по сетчатке и мы его просто не замечаем. (Возможно, что в некотором смысле зрение отключается на период скачка с помощью какой-то сложной нейронной схемы, которая связывает глазодвигательные центры с главным зрительным путем.) Таким образом, процесс обследования зрительного поля при чтении или просто при рассматривании окружающего пространства состоит из серии быстрых скачков с одной точки на другую. Специальная запись движений глаз позволяет наглядно продемонстрировать, насколько незаметны для нас эти скачки. Чтобы записывать движения глаз, берут крошечное зеркальце и прикрепляют его к контактной линзе сбоку — так, чтобы оно не мешало видеть. После этого на зеркальце направляют пучок света, который, отражаясь, создает на экране небольшое световое пятнышко. Есть еще один способ регистрации движений глаз, более современный; он разработан Д. Робинсоном в Институте Уилмера (университета Джонса Гопкинса). При этой методике по краю контактной линзы на глазу испытуемого монтируется крохотная катушка индуктивности, а самого испытуемого помещают между двумя взаимно перпендикулярными обручами величиной с велосипедное колесо, на которых расположены другие катушки индуктивности. Протекание тока в этих катушках ведет к появлению тока в катушке на контактной линзе. После калибровки такой системы можно регистрировать движения глаз с большой точностью. Для самого бедняги испытуемого ни тот, ни другой способ регистрации движений глаз нельзя назвать приятным! В 1957 году советский психофизик А. Л. Ярбус записал движения глаз испытуемых при рассматривании различных изображений, в частности таких, как сцена в лесу и женское лицо (рис. 50). На этих записях периоды фиксации взора представлены точками, а точки соединены линиями, показывающими траекторию движения глаз во время скачков. Даже беглое ознакомление с этими удивительными записями дает много сведений о работе нашей зрительной системы — даже о том, какие предметы и детали окружающего мира нас больше интересуют. Итак, первый факт, противоречащий нашей интуиции, состоит в том, что при зрительном осмотре глаза перескакивают с одной интересной точки на другую и что нельзя осматривать неподвижную сцену плавными движениями глаз. Задача глазодвигательной системы, видимо, заключается не в том, чтобы удерживать изображение на сетчатках неподвижным, а в том, чтобы предотвращать его плавное смещение. Если же движется вся зрительная сцена, как бывает, когда мы смотрим из окна поезда, то мы прослеживаем эту сцену, фиксируя взором какой-нибудь объект и поддерживая его фиксацию путем плавного движения глаз до тех пор, пока объект не выйдет из зоны видимости, после чего делаем скачок и фиксируем новый объект. Такая последовательность движений глаз — плавное прослеживающее движение, скажем вправо, а затем саккада влево — называется нистагмом. Вы сможете сами увидеть такого рода движения, когда окажетесь в поезде или в трамвае: понаблюдайте за движениями глаз ваших попутчиков, когда они будут рассматривать через окно окружающий пейзаж (будьте только осторожны, чтобы ваше внимание не было неверно истолковано!). Управление скачками глаз при рассматривании интересных элементов сцены с целью перевода их изображений в центральную ямку осуществляется из верхних бугорков четверохолмия. Это показал в 1978 году в серии впечатляющих работ П. Шиллер из Массачусетсского технологического института.
 Рис. 50. Испытуемый рассматривает картину, и в это время у него все время регистрируется положение глаз и, следовательно, направление взгляда. Глаза совершают скачок и тотчас останавливаются (в этот момент в записи появляется небольшая точка), затем следует скачок к новому интересному месту. Создается впечатление, что глазу трудно перескочить в те места картины, где нет резких перепадов яркости.
Рис. 50. Испытуемый рассматривает картину, и в это время у него все время регистрируется положение глаз и, следовательно, направление взгляда. Глаза совершают скачок и тотчас останавливаются (в этот момент в записи появляется небольшая точка), затем следует скачок к новому интересному месту. Создается впечатление, что глазу трудно перескочить в те места картины, где нет резких перепадов яркости.
Вторая группа фактов относительно того, как мы видим, еще больше противоречит нашему субъективному впечатлению. Когда мы рассматриваем неподвижную сцену и наш взгляд фиксирует какую-то точку, привлекающую внимание, то эта фиксация не бывает абсолютно неподвижной. Несмотря на все наши попытки жестко фиксировать точку, глаза не остаются в полном покое, а совершают непрерывные микродвижения, называемые микросаккадами. Они совершаются несколько раз в секунду и направлены более или менее случайно, достигая амплитуды 1–2 угловых минут. В 1952 году Л. Риггс и Ф. Рэтлифф из Университета Брауна и Р. Дитчберн и Б. Гинсборг из Редингского университета одновременно и независимо обнаружили, что если изображение на сетчатке искусственно стабилизировать (специальными методами), устранив его смещения относительно сетчатки, то зрительный образ спустя примерно секунду как бы «выцветает» и поле зрения становится совершенно пустым! (Простейший способ стабилизации состоит в том, что точечный источник света прикрепляют к контактной линзе; когда глаза движутся, источник света тоже движется и световое пятно быстро становится невидимым.) Если после стабилизации хотя бы чуть-чуть сместить изображение на сетчатке, световое пятно тотчас появляется снова. Очевидно, микросаккады необходимы для того, чтобы непрерывно видеть неподвижные объекты. Как будто бы Природа, создавая зрительную систему, особенно заботилась о восприятии движения и поэтому постаралась обеспечить нечувствительность клеток к неподвижным объектам, однако потом ей пришлось изобрести микросаккады, для того чтобы сделать и неподвижные объекты видимыми. Можно предположить, что в этом процессе участвуют сложные клетки коры, особо чувствительные к движению стимула, но, вероятно, не участвуют клетки с дирекциональной избирательностью, так как микросаккады явно случайно распределяются по направлениям. С другой стороны, механизм дирекциональной избирательности должен, по-видимому, быть полезным для выявления движений объектов относительно неподвижного фона, сигнализируя о наличии движения и о его направлении. Для того чтобы следить за движущимся объектом на неподвижном фоне, нужно фиксировать объект и перемещать взор вместе с ним. В этом случае изображение всего остального будет передвигаться по сетчатке (такая ситуация в других случаях встречается редко). Перемещение всех деталей неподвижного фона по сетчатке должно приводить к бурной активности клеток коры.
Концы линий как зрительные стимулы
В стриарной коре встречается еще один вид клеток. Обычно для простых и сложных клеток характерна пространственная суммация — чем длиннее стимульная линия, тем лучше ответ. Однако реакция усиливается лишь до тех пор, пока длина линии не достигнет размеров рецептивного поля: дальнейшее удлинение линии не приводит к более энергичному ответу. В отличие от этого у клеток, реагирующих на концы линий (end stopped cells), удлинение линии до известного предела продолжает улучшать ответ, а если линия выходит за этот предел (в одном или в обоих направлениях), то ответ ослабевает (рис. 51, Б). Некоторые клетки, которые мы называем «реагирующими исключительно на конец линии» (completely end stopped cells), вообще не отвечают на предъявление стимула в виде длинной линии. Ту зону, с которой можно вызвать ответ клетки, мы называем зоной активации (или возбуждающей зоной), а зоны, расположенные с одного или с двух концов, — зонами торможения (или тормозными зонами). Таким образом, все рецептивное поле подобной клетки состоит из возбуждающей зоны и тормозной зоны (или зон) по краям. Стимул оптимальной ориентации, активирующий клетку с возбуждающей зоны, вызывает максимальное торможение за пределами этой зоны (с одной или с двух сторон). Это можно показать путем повторной стимуляции возбуждающей зоны линией оптимальной длины с оптимальной ориентацией при одновременном тестировании внешней зоны линиями разной ориентации (как показано на рис. 52).
 Рис. 51. А. Ответ обычной сложной клетки на светлую линию различной длины. Длительность каждой записи — 2 секунды. Как показывает график зависимости ответа от длины линии, реакция данной клетки усиливается, пока линия не достигает длины около 2°, после чего ответ остается неизменным. Б. Для этой клетки, реагирующей на концы линий, ответ возрастает, пока длина линии не достигнет 2°, а затем снижается, так что линия длиной 6° не вызывает никакого ответа.
Рис. 51. А. Ответ обычной сложной клетки на светлую линию различной длины. Длительность каждой записи — 2 секунды. Как показывает график зависимости ответа от длины линии, реакция данной клетки усиливается, пока линия не достигает длины около 2°, после чего ответ остается неизменным. Б. Для этой клетки, реагирующей на концы линий, ответ возрастает, пока длина линии не достигнет 2°, а затем снижается, так что линия длиной 6° не вызывает никакого ответа.
 Рис. 52. Для данной клетки, реагирующей на концы линий, предъявление в средней возбуждающей зоне одной полосы с оптимальной ориентацией приводит к возникновению энергичного ответа. Вовлечение в стимуляцию одной из тормозных зон почти полностью подавляет реакцию клетки; однако, если в этой тормозной зоне стимул имеет иную ориентацию, первоначальный ответ клетки не изменяется. Таким образом, и для возбуждающей зоны, и для тормозных зон оптимальная ориентация одинакова.
Рис. 52. Для данной клетки, реагирующей на концы линий, предъявление в средней возбуждающей зоне одной полосы с оптимальной ориентацией приводит к возникновению энергичного ответа. Вовлечение в стимуляцию одной из тормозных зон почти полностью подавляет реакцию клетки; однако, если в этой тормозной зоне стимул имеет иную ориентацию, первоначальный ответ клетки не изменяется. Таким образом, и для возбуждающей зоны, и для тормозных зон оптимальная ориентация одинакова.
Вначале мы думали, что такие клетки относятся к следующему уровню иерархии корковых нейронов, на уровень выше сложных клеток. Согласно простейшей схеме возможной организации таких клеток, они могли бы иметь один или несколько возбуждающих входов от обычных сложных клеток с рецептивными полями, находящимися в возбуждающей зоне, и тормозные входы от сложных клеток с такой же ориентацией рецептивного поля, расположенных вне зоны возбуждения (эту схему поясняет рис. 53). Согласно другой возможной схеме (рис. 54), клетка имеет возбуждающий вход от клеток с небольшим рецептивным полем (а) и тормозный вход от клеток с большим рецептивным полем (б). Предполагается, что клетки, подающие тормозные сигналы, максимально чувствительны к длинным линиям, но слабо возбуждаются короткими линиями. Эта вторая схема (аналогичная модели клеток, имеющих рецептивные поля с центром и периферией, см. стр. 60) — одна из немногих схем, получивших частичное подтверждение. Чарлз Гилберт из Рокфеллеровского университета в Нью-Йорке установил, что сложные клетки в слое 6 стриарной коры обезьяны имеют как раз те свойства, которые нужны для торможения, предполагаемого в данной схеме. Он также показал, что если инактивировать эти клетки путем локальных инъекций, торасположенные в верхних слоях коры клетки, реагирующие на концы линий, теряют способность отвечать торможением на конец линии.
 Рис. 53. Одна из схем, позволяющих объяснить поведение сложной клетки, реагирующей на концы линий. На такую клетку конвергируют выходы трех обычных сложных клеток; при этом у одной из этих клеток рецептивное поле совпадает по положению с возбуждающей зоной клетки, реагирующей на конец стимула (а), а у двух других клеток рецептивные поля лежат по обе стороны от возбуждающей зоны (б и в). Первая клетка образует на выходе возбуждающий синапс (+), а две другие — тормозные синапсы (–).
Рис. 53. Одна из схем, позволяющих объяснить поведение сложной клетки, реагирующей на концы линий. На такую клетку конвергируют выходы трех обычных сложных клеток; при этом у одной из этих клеток рецептивное поле совпадает по положению с возбуждающей зоной клетки, реагирующей на конец стимула (а), а у двух других клеток рецептивные поля лежат по обе стороны от возбуждающей зоны (б и в). Первая клетка образует на выходе возбуждающий синапс (+), а две другие — тормозные синапсы (–).
 Рис. 54. Альтернативная схема, в которой торможение осуществляет одна клетка с рецептивным полем, соответствующим всей зоне а+б+в на рис. 53. Для того чтобы эта схема работала, мы должны предположить, что тормозная клетка слабо реагирует на короткую линию при стимуляции зоны а, но интенсивно отвечает на длинную линию.
Рис. 54. Альтернативная схема, в которой торможение осуществляет одна клетка с рецептивным полем, соответствующим всей зоне а+б+в на рис. 53. Для того чтобы эта схема работала, мы должны предположить, что тормозная клетка слабо реагирует на короткую линию при стимуляции зоны а, но интенсивно отвечает на длинную линию.
Уже после того как были предложены описанные выше модели, Дж. Генри (Канберра, Австралия) обнаружил простые клетки, реагирующие на конец линии, схема рецептивных полей которых представлена на рис. 55. Схема организации подобных клеток аналогична предложенной выше первой схеме, за исключением того, что входы здесь не от сложных клеток, а от простых. Таким образом, ответы сложных клеток, реагирующих на концы линий, могли бы определяться сочетанием возбуждающего входа от одного набора сложных клеток и тормозного входа от другого набора (как в схемах, представленных на рис. 53 и 54) или же конвергенцией входов от нескольких простых клеток, реагирующих на концы линий.
 Рис. 55. Предполагается, что простая клетка, реагирующая на концы линий, может быть организована за счет конвергенции входных связей от трех обычных простых клеток. (Одна из них, имеющая on-центр, могла бы оказывать возбуждающее действие; две другие могли бы иметь on-центры и возбуждающие выходы или же off-центры и тормозящие выходы.) Вместо этого входные сигналы могли бы поступать непосредственно от клеток с круглыми полями (с центром и периферией) при использовании более сложного варианта схемы, показанной на рис. 44.
Рис. 55. Предполагается, что простая клетка, реагирующая на концы линий, может быть организована за счет конвергенции входных связей от трех обычных простых клеток. (Одна из них, имеющая on-центр, могла бы оказывать возбуждающее действие; две другие могли бы иметь on-центры и возбуждающие выходы или же off-центры и тормозящие выходы.) Вместо этого входные сигналы могли бы поступать непосредственно от клеток с круглыми полями (с центром и периферией) при использовании более сложного варианта схемы, показанной на рис. 44.
Оптимальным стимулом для клетки, настроенной на конец линии, служит отрезок линии определенной длины. Для такой клетки, которая реагирует на границу, причем с одной стороны отвечает только на ее конец, идеальным стимулом будет угол; для клетки, реагирующей на оба конца светлых или темных линий, наилучшим стимулом окажется короткая белая или черная линия, а также такая кривая линия, которая попадает в возбуждающую зону и не попадает из-за своей кривизны в тормозную зону (так будет, если ориентация ее концов отличается не менее чем на 20–30° от ориентации ее средней части; подобный случай показан на рис. 56). Таким образом, клетки, реагирующие на концы, можно рассматривать как клетки, чувствительные к углам, к кривизне или к резким изломам линий.
 Рис. 56. Для клетки, реагирующей на концы линий (вроде представленной на рис. 52), эффективным стимулом может служить искривленная граница.
Рис. 56. Для клетки, реагирующей на концы линий (вроде представленной на рис. 52), эффективным стимулом может служить искривленная граница.
Нейрофизиология одиночных клеток и зрительное восприятие
Сам по себе факт, что какая-то клетка мозга реагирует на зрительные стимулы, еще не означает, что она непосредственно участвует в восприятии. Например, многие нейронные структуры в стволе мозга, относящиеся главным образом к зрительной системе, предназначены только для вспомогательных функций — для управления движениями глаз или сужением зрачка, для фокусировки изображения с помощью хрусталика. Что касается тех клеток, которые я описывал в этой главе, то очевидно, что они имеют самое прямое отношение к зрительному восприятию. Как я уже упоминал вначале, разрушение любого небольшого участка стриарной коры приводит к слепоте в каком-то небольшом участке поля зрения. У обезьян повреждение стриарной коры дает такой же эффект. Однако у кошек дело обстоит сложнее — кошка с удаленной стриарной корой может видеть, хотя и не так хорошо, как раньше. Другие отделы мозга, такие как верхние бугорки четверохолмия, возможно, играют у нее более важную роль в зрительном восприятии, чем у приматов. У таких низших позвоночных, как лягушки и черепахи, нет ничего похожего на нашу кору большого мозга, однако никто не станет утверждать, что они слепы. Теперь мы с достаточной уверенностью можем сказать, что именно любая из описанных выше корковых клеток делает в ответ на стимуляцию изображением видимой сцены. Большинство корковых клеток плохо отвечает на диффузное освещение и хорошо — на линии с нужной ориентацией. Так, при показе фигуры, напоминающей по форме почку (рис. 57), такого рода клетка будет отвечать тогда и только тогда, когда участок границы с определенной ориентацией будет пересекать ее рецептивное поле. Те же клетки, рецептивные поля которых расположены внутри границ фигуры, никак не отреагируют — они будут продолжать давать спонтанный импульсный разряд независимо от присутствия или отсутствия данной фигуры.
 Рис. 57. Насколько вероятно, что клетки в нашем мозгу будут отвечать на те или иные характерные стимулы, такие, например, как изображенная здесь почковидная фигура? Из всего множества клеток зрительной коры реакцию на подобный стимул будет давать только очень небольшая группа клеток.
Рис. 57. Насколько вероятно, что клетки в нашем мозгу будут отвечать на те или иные характерные стимулы, такие, например, как изображенная здесь почковидная фигура? Из всего множества клеток зрительной коры реакцию на подобный стимул будет давать только очень небольшая группа клеток.
Так ведут себя клетки с ориентационной избирательностью. Однако для возбуждения простой клетки недостаточно того, чтобы участок контура соответствовал оптимальной ориентации, — контур должен еще почти точно попадать на границу тормозной и возбуждающей зон рецептивного поля, ведь для ответа нужно, чтобы свет падал на возбуждающую зону, но не распространялся на тормозную. Если хотя бы немного сдвинуть участок контура, не меняя его ориентации, стимуляция данной клетки окажется недостаточной, и теперь уже начнет возбуждаться другая популяция простых клеток. Для сложных клеток условия возбуждения не столь жестки, поскольку популяция клеток, активированная стимулом в какой-то момент времени, не изменится при небольшом сдвиге границ фигуры без изменения их ориентации. Чтобы заметно изменить популяцию возбужденных сложных клеток, нужно достаточно сильно сдвинуть границу — так, чтобы она полностью вышла из рецептивных полей одних клеток и попала в поля других. Таким образом, популяция возбужденных сложных клеток — в отличие от простых — в целом мало меняется при небольших поступательных смещениях объекта. Если, наконец, обратиться к клеткам, реагирующим на концы линий, то и здесь мы обнаружим менее жесткие ограничения на точное положение стимула (хотя каждая данная конфигурация будет активировать гораздо меньше клеток). Для нейронов этого типа ориентация контура должна всегда совпадать с оптимальной ориентацией возбуждающей зоны рецептивного поля, но должна заметно меняться за пределами этой зоны, чтобы торможение не уравновесило возбуждение. Короче говоря, участок контура должен иметь достаточную кривизну или резко прерываться, чтобы были соблюдены все условия для возбуждения данной клетки (см. рис. 56). Эти жесткие требования повышают избирательность реакций коры, так как каждый видимый объект возбуждает лишь очень малую долю клеток, на рецептивные поля которых падает его изображение. Такая специализация клеток, вероятно, продолжает возрастать и при дальнейшем переходе к еще более высоким уровням за пределами стриарной коры. На палочки и колбочки воздействует просто свет как таковой. Ганглиозные клетки сетчатки, клетки НКТ и корковые нейроны с центром и периферией сравнивают соответствующие участки поля зрения с окружающим фоном, поэтому они, вероятно, будут реагировать на любой отрезок контура, попавшего в их рецептивное поле, но не будет давать ответа на общее изменение освещенности сетчатки. Клетки, избирательно чувствительные к ориентации, отмечают не только наличие контура, но и его ориентацию и даже быстроту ее изменения, т.е. кривизну линии. Если такие клетки относятся к категории сложных, то они чувствительны также к движению. Как говорилось в одном из предыдущих разделов книги, возможны два предположения относительно роли чувствительности к движению: возможно, она позволяет легче обращать внимание на движущиеся объекты, а может быть, этот механизм в сочетании с микросаккадами поддерживает реакцию сложных клеток на неподвижные объекты. Мне кажется, что границы между светлым и темным — самый важный компонент наших зрительных восприятий, но, без сомнения, далеко не единственный. Цвет различных объектов, безусловно, помогает при выделении их (хотя нужно сказать, что наши последние работы указывают на меньшую роль цвета при определении формы). При зрительной оценке формы и глубины используются также распределение светотени на поверхности предметов и текстура их поверхности. Хотя клетки, работу которых мы обсуждали, в принципе могли бы участвовать в восприятии светотеневых переходов и текстуры, не следует ожидать, что они смогут четко реагировать на оба этих признака. Еще предстоит выяснить, каков механизм восприятия текстур в зрительной системе. Одна из возможных гипотез состоит в том, что сложные клетки обрабатывают информацию о полутонах и текстурах самостоятельно, без помощи каких бы то ни было специализированных групп клеток. Возможно, что такие стимулы не способны сильно активировать большое число клеток, однако текстурам и полутонам свойственна пространственная протяженность, и они могли бы вызывать реакцию сразу многих клеток, хотя каждая клетка будет отвечать умеренно или слабо. Быть может, вялые ответы множества клеток были бы достаточны для передачи информации на более высокие уровни. Многие люди, в том числе и я, все еще с трудом воспринимают то, что внутренняя часть любой фигуры (например, область внутри почковидного пятна на рис. 57) не является стимулом для клеток нашего мозга и что восприятие однородного внутреннего поля как белого или черного (или окрашенного, как мы увидим в главе 8) зависит только от возбуждения клеток, чувствительных к границам. Логическим доводом в пользу этого служит следующее соображение: если бы при восприятии внутренней области активировались клетки, рецептивные поля которых лежат в ее пределах, то она должна была бы, наоборот, восприниматься как неоднородная! Таким образом, если мы видим все занятое фигурой пространство как равномерно черное, белое, серое или зеленое, то клетки с рецептивными полями внутри границ фигуры не имеют к этому никакого отношения. С этой мыслью трудно смириться, не так ли? Однако с точки зрения инженера, конструирующего машину для кодирования формы, такой механизм, я думаю, был бы очень подходящим. Единственная информация, которую необходимо в этом случае получать, — это информация о внешнем контуре фигуры; анализ ее внутренности становится излишним. Нужно ли удивляться, что эволюция мозга пошла по пути, позволяющему перерабатывать информацию с помощью минимального количества клеток? Услышав о свойствах простых и сложных клеток, люди часто делают вывод, что для полного анализа всех мельчайших элементов зрительного поля, включая темные и светлые линии и края различных участков во всех возможных ориентациях, требуется астрономическое число клеток. Разумеется, так оно и есть. Но все дело в том, что кора как раз и содержит астрономическое число клеток. Сегодня мы уже можем сказать, как работают клетки в данном участке мозга, по крайней мере как они отвечают на множество простых зрительных стимулов, встречающихся в повседневной жизни. Я подозреваю, что в стриарной коре нет двух клеток, которые работали бы совершенно одинаково. Действительно, всякий раз, когда с помощью микроэлектрода удавалось одновременно регистрировать активность двух клеток, оказывалось, что эти клетки хоть немного, но отличаются друг от друга — по положению рецептивных полей, дирекциональной чувствительности, интенсивности ответов или каким-то другим параметрам. Короче говоря, складывается впечатление, что в данном отделе мозга если и есть избыточность, то она невелика. Можем ли мы быть уверены в том, что описанные клетки на самом деле предназначены именно для выявления отрезков прямых линий, а не каких-то других стимулов? Нельзя сказать, что мы и другие исследователи не пытались применять множество других стимулов, включая лица, карту мира и пассы руками. Как показывает опыт, было бы глупо думать, что мы исчерпали все возможности. В начале 1960-х годов, когда мы удовлетворились результатами, полученными с клетками стриарной коры, и решили перейти (и даже фактически уже перешли) к следующей области, нам случайно удалось записать реакцию одной слабо отвечавшей клетки стриарной коры. Однако, сделав светлую линию более короткой, мы убедились, что эта клетка способна давать весьма энергичную реакцию. Именно тогда мы наткнулись на класс клеток, отвечающих на концы линий. После этого мы еще почти двадцать лет работали с корковыми клетками обезьян, прежде чем обнаружили «пузырьки» — скопления клеток, специфически реагирующих на цвет (они описаны в главе 8). Сделав эти оговорки, я должен добавить, что некоторые из свойств, обнаруженных у клеток стриарной зоны, например ориентационная чувствительность, являются, несомненно, истинными свойствами этих клеток. В пользу этого можно привести много дополнительных данных, например данные по функциональной анатомии, описанные в главе 5.
Бинокулярная конвергенция
До сих пор я почти не упоминал о существовании двух глаз. Очевидный интерес представляет вопрос: получает ли та или иная корковая клетка входные сигналы от обоих глаз, и если да, то одинаковы ли эти входы в количественном и качественном отношении? Для того чтобы получить ответ, мы должны вернуться на некоторое время к наружному коленчатому телу (НКТ) и выяснить, имеют ли какие-то из его клеток входы от обоих глаз. НКТ — это самый низкий уровень, на котором уже возможно было бы объединение сигналов от двух глаз на одной клетке. Однако возможность эта, по-видимому, здесь не реализована — два разных пучка входных волокон распределяются по разным клеточным слоям, между которыми нет или почти нет взаимодействия. Как можно было бы ожидать, учитывая раздельность проекций от двух глаз, отдельная клетка НКТ должна отвечать на стимуляцию одного какого-то глаза и совсем не реагировать на стимуляцию другого. Судя по результатам некоторых экспериментов, стимулы, подаваемые на «чужой» для данной клетки НКТ глаз, могут оказывать слабое воздействие на реакцию, вызываемую со «своего» глаза. Однако практически можно считать, что каждая клетка находится под контролем только одного глаза. Интуитивно ясно, что пути, идущие от каждого глаза, должны рано или поздно сойтись вместе, так как мы, глядя на что-либо, видим одну целостную картину. Тем не менее повседневный опыт говорит нам, что если закрыть один глаз, то большого изменения не произойдет — предметы будут казаться такими же четкими, такими же реальными и такими же яркими. Разумеется, общее поле зрения для двух глаз будет несколько шире, поскольку каждый глаз видит на своей стороне бо́льшую область пространства, чем другой глаз, хотя эта разница составляет всего лишь около 20–30°. Существенное отличие бинокулярного зрения от монокулярного состоит в ощущении глубины, о чем пойдет речь в главе 7. В коре обезьяны те клетки, которые получают входные сигналы от НКТ и имеют рецептивные поля с круговой симметрией, сходны с клетками НКТ в том, что они тоже монокулярные. На этом корковом уровне мы находим примерно одинаковое число клеток, возбуждающихся от левого и от правого глаза, по крайней мере в тех участках коры, которые обслуживают область поля зрения в радиусе около 20° от центральной точки фиксации. Однако на следующем уровне коры выявляются уже бинокулярные клетки, простые и сложные, причем у макаков больше половины таких клеток могут реагировать независимо на сигналы от каждого из глаз. Как только мы нашли бинокулярную клетку, можно тщательно сравнить ее рецептивные поля в обеих сетчатках. Сначала закрываем правый глаз животного и картируем рецептивное поле клетки в левом глазу, отмечая его точное положение на экране или на сетчатке, а также его сложность, ориентацию и расположение возбуждающих и тормозных зон; выясняем также, простая это клетка или сложная, исследуем ее способность реагировать на конец линии и дирекциональную чувствительность; после этого закрываем левый глаз животного, открываем правый и повторяем всю процедуру измерений сначала. Оказалось, что у большинства бинокулярных клеток все свойства, выявляемые в опытах с левым глазом, обнаруживаются и при стимуляции правого глаза — то же положение на сетчатке, та же дирекциональная чувствительность и т.д. Это позволяет заключить, что все связи, идущие к данной клетке от левого глаза, совпадают по структуре со связями, идущими от правого глаза. Говоря о таком дублировании связей, нужно сделать одно уточнение. Если, определив для клетки оптимальный стимул, его положение, ориентацию, направление движения и т.д., мы сравним ее ответы при стимуляции одного глаза и при стимуляции другого, интенсивность реакции не всегда окажется одинаковой. Некоторые клетки действительно одинаково хорошо активируются от обоих глаз, однако другие явно дают более сильный разряд при стимуляции определенного глаза. В целом, за исключением той части корковых клеток, которые обслуживают периферию поля зрения, мы не находим никакого особого преимущества того или другого глаза — в каждом полушарии число клеток, лучше активируемых с противоположной стороны (от контралатерального глаза) и с той же стороны (от ипсилатерального глаза), примерно одинаково. При этом встречаются все степени относительного доминирования глаза, начиная от клеток, возбуждаемых исключительно с левого глаза, и кончая клетками, отвечающими только на стимуляцию правого глаза. Теперь можно оценить численность различных популяций клеток. Разобьем все изученные клетки, скажем 1000 штук, произвольно на семь классов по относительной эффективности воздействия на них того или другого глаза. Затем подсчитаем число клеток в каждом классе. На рис. 58 представлены соответствующие гистограммы для кошки и макака. Здесь сразу видны сходства и различия в распределении клеток у этих животных. Видно, что у обоих видов бинокулярные клетки встречаются достаточно часто, причем среди клеток с односторонним доминированием хорошо представлены оба глаза (у макака примерно поровну), что у кошек бинокулярных клеток очень много, что у макаков численность монокулярных и бинокулярных клеток примерно одинакова, причем у бинокулярных клеток зачастую сильно выражено доминирование одного глаза (группы 2 и 5), и что реже всего встречаются клетки, одинаково хорошо активируемые с любого глаза.
 Рис. 58. Изучая распределение нейронов по глазодоминантности, мы исследовали сотни клеток и относили каждую из них к одной из семи произвольно выделенных групп. Клетки группы 1 определяются как клетки, на которые влияет только контралатеральный глаз, т.е. глаз, находящийся на противоположной стороне тела; клетки группы 2 реагируют на стимуляцию обоих глаз, однако явно предпочитают сигналы от контралатерального глаза; и т.д.
Рис. 58. Изучая распределение нейронов по глазодоминантности, мы исследовали сотни клеток и относили каждую из них к одной из семи произвольно выделенных групп. Клетки группы 1 определяются как клетки, на которые влияет только контралатеральный глаз, т.е. глаз, находящийся на противоположной стороне тела; клетки группы 2 реагируют на стимуляцию обоих глаз, однако явно предпочитают сигналы от контралатерального глаза; и т.д.
 Рис. 59. Регистрирующий электрод оказался достаточно близко к трем клеткам, чтобы отводить импульсы от всех трех. Ответы разных клеток можно различить по амплитуде и форме импульсов. На рисунке показаны реакции на стимулы, предъявленные одному глазу и обоим глазам. Клетки 1 и 2 должны быть отнесены к группе 4, так как они почти одинаково отвечают на стимуляцию того и другого глаза. Клетка 3 отвечает только на одновременную стимуляцию обоих глаз; можно сказать лишь то, что она не принадлежит ни к группе 1, ни к группе 7.
Рис. 59. Регистрирующий электрод оказался достаточно близко к трем клеткам, чтобы отводить импульсы от всех трех. Ответы разных клеток можно различить по амплитуде и форме импульсов. На рисунке показаны реакции на стимулы, предъявленные одному глазу и обоим глазам. Клетки 1 и 2 должны быть отнесены к группе 4, так как они почти одинаково отвечают на стимуляцию того и другого глаза. Клетка 3 отвечает только на одновременную стимуляцию обоих глаз; можно сказать лишь то, что она не принадлежит ни к группе 1, ни к группе 7.
Теперь мы можем продвинуться дальше и задать следующий вопрос: отвечают ли бинокулярные клетки при стимуляции обоих глаз лучше, чем при стимуляции одного глаза? Оказывается, многие клетки именно таковы — от одного глаза они активируются слабо или не возбуждаются совсем, а от двух глаз дают сильный импульсный разряд, особенно в том случае, когда оба глаза стимулируются одновременно и совершенно одинаково. На рис. 59 показаны записи ответов трех клеток (1, 2 и 3), демонстрирующих выраженную синергию. Одна из этих клеток вообще не отвечает на стимуляцию только одного глаза, так что мы вообще не смогли бы ее обнаружить, если бы не стимулировали оба глаза одновременно. Эффект синергии у многих клеток выражен слабо или не отмечается вовсе — такие клетки отвечают на стимуляцию обоих глаз примерно так же, как и на стимуляцию каждого глаза по отдельности. Подобные связи одиночных клеток с двумя глазами еще раз указывают на высокую степень специфичности соединений в мозгу. Мало того, что входные системы связей данной клетки позволяют ей отвечать только на линию определенной ориентации и лишь на одно направление движения, — оказывается к тому же, что эти системы представлены двумя копиями, по одной от каждого глаза. Но и этого еще мало: как мы узнаем из главы 9, большинство связей, по-видимому, должно быть сформировано и готово к работе уже к моменту рождения животного. Все это поистине удивительно.
5. Архитектура зрительной коры
По сравнению с наружными коленчатыми телами (НКТ) и сетчаткой первичная зрительная кора, или стриарная кора, — структура гораздо более сложная. Как мы уже видели, резкое увеличение структурной сложности отделов мозга сопровождается таким же усложнением физиологической организации. В стриарной коре мы находим большее разнообразие функциональных типов клеток. Нейроны стриарной коры отвечают на более сложные стимулы, т.е. стимулы с большим числом параметров, причем эти параметры должны быть вполне определенными. Если при исследовании клеток сетчатки и НКТ нам достаточно было изменять только местоположение и размеры стимула в виде простого пятна, то теперь мы внезапно столкнулись с необходимостью учитывать такие параметры, как ориентация линии, направление движения, длина линии и ее кривизна, а также выбор глаза, на который подается стимул. Какая связь существует между этими параметрами и структурной организацией коры (если она вообще есть)? Для того чтобы подойти к рассмотрению этого вопроса, сначала необходимо сообщить кое-что о строении стриарной коры.Анатомия зрительной коры
Кора представляет собой слой нервной ткани толщиной около 2 мм, который почти полностью покрывает большие полушария головного мозга. Площадь поверхности коры у человека составляет больше квадратного фута (около 900 см2). Общая площадь коры у макаков примерно в 10 раз меньше, чем у человека. Уже более столетия известно, что кора полушарий подразделяется на множество различных корковых полей. Среди всех этих полей первой удалось выделить зону первичной зрительной коры, которая на поперечных срезах выглядит слоистой или полосатой (отсюда и ее давнее название — стриарная кора: лат. stria — полоса). Было время, когда все устремления нейроморфологов сводились к тому, чтобы выявить как можно больше корковых полей на основании подчас весьма тонких гистологических различий. В одной из принятых систем такой классификации стриарной коре был присвоен номер 17. По одной из новейших оценок (Д. ван Эссен, Калифорнийский технологический институт) у макаков первичная зрительная кора занимает около 1200 мм2 (немногим меньше одной трети кредитной карточки); это примерно 15% всей площади коры, т.е. довольно существенная ее часть.
 Рис. 60. На этом срезе видны колонки глазодоминантности в коре левого полушария мозга макака. Срез сделан перпендикулярно поверхности коры в направлении слева направо. Прослеживая поверхность коры в направлении слева направо (см. верхнюю часть фотографии), мы увидим, что она образует изгиб и глубокую складку, проходящую справа налево. Радиоактивная аминокислота, введенная путем инъекции в левый глаз животного, транспортировалась через НКТ в слой 4C коры и сосредоточилась во множестве отдельных участков толщиной каждый в полмиллиметра, которые выделяются как яркие полоски на общем темном фоне. (Сплошной светлый листок в середине — это белое вещество, состоящее из волокон от коленчатого тела.)
Рис. 60. На этом срезе видны колонки глазодоминантности в коре левого полушария мозга макака. Срез сделан перпендикулярно поверхности коры в направлении слева направо. Прослеживая поверхность коры в направлении слева направо (см. верхнюю часть фотографии), мы увидим, что она образует изгиб и глубокую складку, проходящую справа налево. Радиоактивная аминокислота, введенная путем инъекции в левый глаз животного, транспортировалась через НКТ в слой 4C коры и сосредоточилась во множестве отдельных участков толщиной каждый в полмиллиметра, которые выделяются как яркие полоски на общем темном фоне. (Сплошной светлый листок в середине — это белое вещество, состоящее из волокон от коленчатого тела.)
 Рис. 61. Здесь видна значительная часть коры правого полушария головного мозга, открытая под местной анестезией для нейрохирургической операции по поводу эпилепсии. Больной находился в полном сознании. Операцию проводил хирург д-р У. Фейндел в Монреальском неврологическом институте. Участок скальпа был отвернут и соответствующий кусок черепа удален (временно, до окончания операции). На снимке видны борозды, извилины, крупные вены пурпурного оттенка и более мелкие (красного цвета) артерии. Общий розоватый оттенок открытой части мозга объясняется наличием мелкой сети разветвлений этих сосудов. Нижняя треть обнаженного участка — височная доля. Выше горизонтально идущих крупных вен расположены части теменной доли (слева) и лобной доли (справа). У самого левого края можно видеть полоску затылочной доли. Операция проводилась с целью лечения определенной формы эпилепсии и заключалась в удалении пораженной части мозга. Такая операция допускается лишь в том случае, если не может привести к нарушению произвольных движений или речи. Для того чтобы избежать этого, нейрохирург путем раздражения электрическим током идентифицирует области, связанные с речью, движениями и чувствительностью, выясняя, какие при этом возникают двигательные эффекты, ощущения в различных частях тела и нарушения речи. Ясно, что такие испытания были бы невозможны, если бы больной не был в сознании. Места, где производилось раздражение, отмечены кусочками стерильной бумаги с цифрами. При раздражении отмечались, например, такие результаты: 1) ощущение покалывания в большом пальце левой руки; 2) покалывание в безымянном пальце левой руки; 3) покалывание в безымянном и среднем пальцах левой руки; 4) сгибание пальцев левой руки и запястья. При раздражении участков с номерами 8 и 13 возникали более сложные явления, сходные с образами памяти, что бывает у некоторых больных эпилепсией при раздражении височной доли.
Рис. 61. Здесь видна значительная часть коры правого полушария головного мозга, открытая под местной анестезией для нейрохирургической операции по поводу эпилепсии. Больной находился в полном сознании. Операцию проводил хирург д-р У. Фейндел в Монреальском неврологическом институте. Участок скальпа был отвернут и соответствующий кусок черепа удален (временно, до окончания операции). На снимке видны борозды, извилины, крупные вены пурпурного оттенка и более мелкие (красного цвета) артерии. Общий розоватый оттенок открытой части мозга объясняется наличием мелкой сети разветвлений этих сосудов. Нижняя треть обнаженного участка — височная доля. Выше горизонтально идущих крупных вен расположены части теменной доли (слева) и лобной доли (справа). У самого левого края можно видеть полоску затылочной доли. Операция проводилась с целью лечения определенной формы эпилепсии и заключалась в удалении пораженной части мозга. Такая операция допускается лишь в том случае, если не может привести к нарушению произвольных движений или речи. Для того чтобы избежать этого, нейрохирург путем раздражения электрическим током идентифицирует области, связанные с речью, движениями и чувствительностью, выясняя, какие при этом возникают двигательные эффекты, ощущения в различных частях тела и нарушения речи. Ясно, что такие испытания были бы невозможны, если бы больной не был в сознании. Места, где производилось раздражение, отмечены кусочками стерильной бумаги с цифрами. При раздражении отмечались, например, такие результаты: 1) ощущение покалывания в большом пальце левой руки; 2) покалывание в безымянном пальце левой руки; 3) покалывание в безымянном и среднем пальцах левой руки; 4) сгибание пальцев левой руки и запястья. При раздражении участков с номерами 8 и 13 возникали более сложные явления, сходные с образами памяти, что бывает у некоторых больных эпилепсией при раздражении височной доли.
На рис. 62 представлена фотография мозга макака (вид сзади). Черепная коробка была удалена, а мозг с целью консервации перфузировали разбавленным раствором формальдегида, что придало ему желтую окраску. Обычно на поверхности коры бросается в глаза сеть кровеносных сосудов, однако здесь они находятся в спавшемся состоянии и поэтому незаметны. На этой фотографии видна главным образом поверхность затылочной доли коры, т.е. та область, которая имеет прямое отношение к зрению и включает не только стриарную кору, но также и одну-две дюжины (а может быть, и больше) престриарных зон. Для того чтобы поместить лоскут нервной ткани толщиной 0,5 мм, равный по площади большой каталожной карточке, в коробку величиной с череп макака, пришлось бы скомкать этот лоскут, наподобие того как мы комкаем лист бумаги, прежде чем выбросить его в мусорную корзину. В результате аналогичного сжимания в коре мозга образуются щели, или борозды, между которыми проходят выпуклые участки, называемые извилинами.
 Рис. 62. Мозг макака (вид сзади). Показана затылочная доля и часть стриарной коры, выходящая на поверхность этой доли (ниже пунктирной линии).
Рис. 62. Мозг макака (вид сзади). Показана затылочная доля и часть стриарной коры, выходящая на поверхность этой доли (ниже пунктирной линии).
Область на фотографии, лежащая позади (ниже) пунктирной линии, — это открытая, видимая часть стриарной коры. Хотя стриарная кора занимает бо́льшую часть поверхности затылочной доли, на фото мы можем увидеть только часть ее (0,3–0,5 всей площади); остальная часть скрыта в борозде. Стриарная кора (поле 17) имеет множество выходных волокон, значительная часть которых идет в следующую корковую область — зрительное поле 2, часто также называемое полем 18, так как оно примыкает к полю 17. Поле 18 представляет собой полоску коры шириной примерно 6–8 мм, почти полностью окружающую поле 17. На фото (рис. 62) можно видеть только часть поля 18, расположенную над пунктирной линией, которая соответствует границе между полями 17 и 18; бо́льшая часть поля 18 уходит вниз, в глубокую борозду, расположенную как раз перед пунктирной линией. Поле 17 упорядоченно, точка в точку, проецируется на поле 18, а последнее в свою очередь образует проекции по крайней мере в трех затылочных зонах величиной каждая с почтовый штемпель. Эти зоны известны как поле MT (сокращение от medialis temporalis, т.е. срединное височное) и зрительные поля 3 и 4 (часто обозначаемые как V3 и V4). Как правило, каждая корковая зона имеет проекции в нескольких других зонах, расположенных на более высоком уровне. Кроме того, для каждой зоны коры можно найти обратную проекцию — в ту зону или те зоны, откуда идут входные волокна. И как будто стремясь к еще большей сложности, каждая из этих зон образует проекции в глубинных структурах мозга, например в верхних бугорках четверохолмия и в различных участках таламуса (сложного клеточного образования величиной с мяч для гольфа, небольшую часть которого составляет НКТ). И все эти зрительные зоны получают входные сигналы от разных участков таламуса — так же, как НКТ проецируется в первичную зрительную кору, так различные области таламуса образуют проекции в других корковых зонах. На той же фотографии буквой X обозначен тот участок поля 17, который получает входные сигналы от центральных ямок обеих сетчаток. Если двигаться по левому полушарию из точки X в направлении, указанном стрелкой, то в поле зрения это будет соответствовать движению объекта по горизонтали от точки фиксации. Если же, выйдя снова из точки X, двигаться прямо вдоль границы между полями 17 и 18, то это будет соответствовать движению объекта от точки фиксации вниз, а перемещение в обратном направлении — движению объекта вверх. Самой стрелке в поле зрения соответствует горизонтальная линия длиной около 6°. Области поля зрения, расположенные дальше 9° от центра взора, отображаются в ту часть поля 17, которая лежит в складке коркового слоя под наружной поверхностью мозга, параллельно ей. Для того чтобы посмотреть, как выглядит кора в поперечном разрезе, мы вырезали участок зрительной коры из правой половины мозга, изображенного на рис. 62. Полученный срез был обработан крезиловым фиолетовым, который окрашивает тела нейронов в синий цвет, но не окрашивает аксоны и дендриты. На этой микрофотографии, сделанной при малом увеличении, нельзя различить отдельные клетки, но можно видеть темные слои плотно упакованных клеток и более светлые слои, где клеток гораздо меньше. Под наружной поверхностью коры видна сложная структура из складок, напоминающая по форме гриб. В действительности эта структура и поверхностная часть коры без перерыва переходят друг в друга. Светлоокрашенное пространство — это белое вещество, оно заполняет области между наружными частями коры и ее глубже расположенными складками. Белое вещество состоит в основном из миелинизированных нервных волокон, которые не окрашиваются красителем. Часть коры, содержащая тела нервных клеток, их аксоны и дендриты, а также синапсы, образует серое вещество.
 Рис. 63. Срез затылочной доли мозга (из участка, показанного на рис. 62): он соответствует левой стенке канавки, вырезанной в правом полушарии). Буквой а отмечена точка, лежащая на уровне стрелки X–>. Окраска по Нисслю позволяет видеть только тела клеток, но они настолько малы, что выглядят как точки. Более темные участки вверху и расположенная ниже грибовидная складка соответствуют стриарной коре. Тремя буквами d указаны границы между полями 17 и 18.
Рис. 63. Срез затылочной доли мозга (из участка, показанного на рис. 62): он соответствует левой стенке канавки, вырезанной в правом полушарии). Буквой а отмечена точка, лежащая на уровне стрелки X–>. Окраска по Нисслю позволяет видеть только тела клеток, но они настолько малы, что выглядят как точки. Более темные участки вверху и расположенная ниже грибовидная складка соответствуют стриарной коре. Тремя буквами d указаны границы между полями 17 и 18.
По сложности своей слоистой структуры поле 17 превосходит все другие зоны коры. Эту разницу можно заметить даже при малом увеличении (рис. 63), если сравнить поле 17 и соседнюю область справа — поле 18 (граница между ними отмечена буквой г). Еще одна деталь: если сравнить на рис. 63 области а, б и в, куда проецируются разные участки поля зрения (расстояния их от точки фиксации <5, 6 и 80–90 градусов соответственно), мы не заметим существенного различия ни в толщине коры, ни в ее слоистой структуре. Как выяснилось, такая однородность имеет важное значение. К этому вопросу я еще вернусь в этой главе.
Слои зрительной коры
На рис. 64 представлен небольшой участок поля 17 при более сильном увеличении. Здесь уже можно различить в виде точек тела отдельных клеток и получить некоторое представление об их размерах, количестве и распределении. Видимая картина слоев частично определяется интенсивностью окрашивания клеток и частично — плотностью их упаковки. Слои 4C и 6 — самые плотные и темные. В слоях 1, 4B и 5 клетки расположены наиболее рыхло. Слой 1 едва ли содержит хотя бы одну нервную клетку, но здесь множество аксонов, дендритов и синапсов. Для того чтобы показать, что разные слои включают разные типы клеток, нужно применить какой-либо вариант методики, изобретенной в 1900 году К. Гольджи. При окраске по Гольджи выявляются лишь немногие клетки, зато на срезах клетка видна вся целиком, включая ее аксон и дендриты. Два главных класса корковых клеток — это пирамидные клетки, имеющиеся во всех слоях коры, кроме слоев 1 и 4, и звездчатые клетки, которые есть во всех слоях. Примеры пирамидной и звездчатой клеток представлены на рис. 4. Типичное распределение пирамидных клеток в коре показано на рисунке Рамон-и-Кахала, помещенном в его книге «Гистология» (рис. 65); здесь изображена лишь небольшая доля всех пирамидных клеток, имеющихся в этом участке, наверное около 1 процента.
 Рис. 64. Поперечный срез стриарной коры при большем увеличении. Видны клетки, расположенные слоями. Слои 2 и 3 невозможно отграничить друг от друга; слой 4A очень тонок. Толстый светлый слой внизу — белое вещество.
Рис. 64. Поперечный срез стриарной коры при большем увеличении. Видны клетки, расположенные слоями. Слои 2 и 3 невозможно отграничить друг от друга; слой 4A очень тонок. Толстый светлый слой внизу — белое вещество.
 Рис. 65. Окрашенный по Гольджи срез слоев 1, 2 и 3 зрительной коры младенца в возрасте нескольких дней. Черные треугольнички — тела клеток, от которых отходят апикальные дендриты, поднимающиеся вверх и разветвляющиеся в слое 1, базальные дендриты, идущие в боковых направлениях, и один тонкий аксон, направляющийся прямо вниз.
Рис. 65. Окрашенный по Гольджи срез слоев 1, 2 и 3 зрительной коры младенца в возрасте нескольких дней. Черные треугольнички — тела клеток, от которых отходят апикальные дендриты, поднимающиеся вверх и разветвляющиеся в слое 1, базальные дендриты, идущие в боковых направлениях, и один тонкий аксон, направляющийся прямо вниз.
Волокна, идущие из НКТ, приходят в кору из белого вещества мозга. Пройдя в диагональном направлении, большая часть волокон доходит до слоя 4C, многократно разветвляется и, наконец, достигает конечного пункта, образуя синапсы со звездчатыми клетками, которые в основном заполняют этот слой. Аксоны, выходящие их двух вентральных (крупноклеточных) слоев НКТ, оканчиваются в верхней половине слоя 4C, называемой подслоем 4Cα, a те, которые выходят из четырех дорсальных (мелкоклеточных) слоев НКТ, оканчиваются в нижней половине слоя 4C (подслой 4Cβ). Как видно из схемы на рис. 66, эти два подслоя образуют разные проекции в верхних слоях — 4Cα посылает сигналы в 4B, а 4Cβ — в слои 2 и 3. Последние в свою очередь тоже различаются по своим проекциям. Заметное различие в нервных путях, выходящих из двух групп слоев НКТ, — это одно из многих оснований предполагать, что с этими путями связаны две разные системы. У большинства пирамидных клеток в слоях 2, 3, 4B, 5 и 6 аксоны выходят из коры, однако у этих аксонов есть ответвления (коллатерали), которые образуют локальные связи и таким образом помогают распределить информацию по всей толще коры.
 Рис. 66. Главные пути от HKT к стриарной коре и от стриарной коры к другим областям мозга. В правой части схемы отмечены зоны с наибольшей плотностью окраски по Нисслю (для сравнения с рис. 64).
Рис. 66. Главные пути от HKT к стриарной коре и от стриарной коры к другим областям мозга. В правой части схемы отмечены зоны с наибольшей плотностью окраски по Нисслю (для сравнения с рис. 64).
Корковые слои различаются не только своими входами и внутренними связями, но также и адресами тех структур, куда они проецируются. Все они, кроме слоев 1, 4A и 4C, имеют выходные волокна, уходящие за пределы коры. Верхние слои 2 и 3, а также слой 4B посылают сигналы главным образом в другие области коры, тогда как нижние слои проецируются на подкорковые структуры: из слоя 5 выходные волокна идут в средний мозг, в верхние бугорки четверохолмия, а из слоя 6 волокна направляются обратно в НКТ. Хотя уже почти столетие известен тот факт, что выходные волокна НКТ идут в основном в корковый слой 4, мы ничего не знали о различиях между выходами разных слоев коры, пора в 1969 году японский исследователь К. Тояма не обнаружил впервые эти различия в физиологических экспериментах. С тех пор эти данные были многократно подтверждены морфологическими методами. Рамон-и-Кахал был первым, кто понял, насколько коротки внутренние связи в коре. Как уже говорилось, наиболее многочисленные связи идут вверх и вниз, тесно объединяя разные слои. Диагональные и боковые (горизонтальные) связи обычно имеют длину от 1 до 2 мм, хотя некоторые связи прослеживаются на расстояние 4–5 мм. Такое ограничение «горизонтальной» передачи информации в коре приводит к важным последствиям. Если входы организованы по топографическому принципу (в случае зрительной системы — в соответствии с положением отображаемой точки на сетчатке или в поле зрения), то этот же принцип должен сохраняться и в организации выходов. Какие бы функции ни выполняла кора, проводимый анализ должен быть локальным. Информация о любом небольшом участке поля зрения поступает в какой-то небольшой участок коры, где подвергается преобразованию, анализу, переработке (назовите это как угодно) и передается куда-нибудь для дальнейшей обработки независимо от того, что происходит с информацией о соседнем участке. Таким образом, изображение зрительной сцены анализируется как бы мозаично. Поэтому первичная зрительная кора не может быть тем местом, где распознаются, воспринимаются или подвергаются другой обработке целые объекты, например лодки, шляпы, лица, т.е. не может быть субстратом процессов «восприятия». Разумеется, столь общий вывод вряд ли правомерно делать на основе одних только анатомических данных: ведь информация могла бы передаваться на большие расстояния вдоль коры и по принципу эстафеты, с отдельными этапами по одному миллиметру. Однако, регистрируя активность клеток при одновременной стимуляции сетчатки, можно показать, что дело обстоит не так: выясняется, что все клетки в данном небольшом участке имеют маленькие рецептивные поля и что соседние клетки всегда имеют рецептивные поля, расположенные почти в одном и том же месте. Нет никаких физиологических данных, которые указывали бы на то, что какая-либо клетка в первичной зрительной коре обезьяны соединена с какой-то другой клеткой на расстоянии больше 2–3 миллиметров. На «мозаичный» характер обработки информации давно уже указывают и данные клинической неврологии. После небольших инсультов, опухолей или ранений первичной зрительной коры может наступать полная слепота в небольших, четко ограниченных участках поля зрения. При этом во всех остальных участках сохраняется нормальное зрение. Если бы каждая клетка была в какой-то мере связана со всеми другими, то вместо этого можно было бы ожидать некоторого общего, нелокального ухудшения зрения. Отойдя несколько в сторону от нашей темы, следует заметить, что больные с такими поражениями могут и не осознавать наличия у них дефекта поля зрения, особенно в тех случаях, когда не затронута та часть коры, где находится проекция центральной ямки сетчатки, т.е. точки фиксации взора. Во всяком случае, такие больные не видят в поле зрения ни сплошь черных или сплошь серых островков, ни каких-либо других дефектов. Даже если разрушена вся затылочная доля на одной стороне и человек ничего не видит в противоположной половине поля зрения, то у него не возникает впечатления, что с этой стороны зрительный мир от него закрыт. У меня самого иногда бывают приступы мигрени (к счастью, без головной боли) с кратковременной слепотой, нередко в значительной части одного из полуполей зрения. Если спросить, что именно я в этой области вижу, я могу только сказать, что не вижу буквально ничего — ни белого, ни серого, ни черного, просто ничего. Еще одна любопытная особенность, наблюдаемая у больных с островком локальной слепоты, или скотомой, — это феномен, известный под названием «заполнения». Если человек со скотомой будет смотреть на линию, проходящую через дефектный участок поля зрения, он не заметит никакого разрыва — линия кажется совершенно непрерывной. Можно продемонстрировать аналогичное явление, воспользовавшись тем, что и в нормальном глазу имеется слепое пятно. Это то место, где из глаза выходит зрительный нерв. Слепое пятно представляет собой овальный участок величиной около двух миллиметров, где нет ни палочек, ни колбочек. Отыскать слепое пятно можно с помощью небольшого бумажного диска — метод настолько прост, что доступен каждому. Сначала закройте один глаз, например левый. Держа его закрытым, фиксируйте другим глазом какой-нибудь маленький предмет в комнате. Теперь возьмите в руку бумажный диск и отведите руку на всю длину, поместив ее точно на линии, соединяющей глаз с предметом. Затем медленно сдвигайте руку вправо точно по горизонтали (наличие темного фона облегчает эксперимент). Бумажный диск исчезнет примерно в тот момент, когда смещение его достигнет 18°. А теперь если взять какой-нибудь стержень и поместить его так, чтобы он пересекал слепое пятно, то он будет выглядеть как целостный объект, не имеющий никаких разрывов. Область, занимаемая слепым пятном, аналогична скотоме — вы не заметите ее существования до тех пор, пока не проделаете описанный выше опыт. Вы не видите здесь ни черного, ни серого, ни чего-либо иного, вы просто ничего не видите. Сходным образом, если смотреть на большой лист белой бумаги, то в коре будут возбуждаться только те клетки, рецептивные поля которых попадают на край этого листа (поскольку корковые клетки имеют склонность игнорировать равномерно распределенный свет). В этом случае прекращение активности клеток, рецептивные поля которых целиком лежат в пределах листа бумаги, должно приводить к исчезновению различий яркостив этой области поля зрения. Островок слепоты в данном случае не должен быть виден — и он не виден. Мы не можем увидеть свое слепое пятно в виде черного отверстия в листе белой бумаги, на которую смотрим. Феномен заполнения, проявляющийся в опыте с листом бумаги, должен убедить каждого в том, что, исходя из одних только интуитивных представлений, мы не сможем понять, как работает на самом деле мозг.
Архитектура коры
Теперь мы можем вернуться к исходному вопросу: как связаны между собой физиологические свойства корковых клеток и их структурная организация? Можно еще больше заострить этот вопрос: зная, что корковые клетки могут различаться по положению их рецептивных полей, «сложности», предпочитаемой ориентации стимула, глазодоминантности, по оптимальному направлению движения стимула и оптимальной длине стимульной линии, правомерно ли ожидать, что соседние клетки сходны по некоторым из этих параметров или даже по всем? Или же клетки с разными свойствами просто рассыпаны по всей коре случайным образом, без всякой связи с их физиологическими свойствами? Если изучать анатомию коры невооруженным глазом или под микроскопом, это мало что даст. На поперечных срезах можно увидеть отчетливые различия между отдельными слоями коры. Однако если прослеживать на поперечном срезе тот же слой по длине или же изучать срезы одного слоя, сделанные параллельно границе слоев, то мы увидим один лишь серый однородный материал. Хотя такая однородность может указывать на случайное распределение клеток, мы знаем, что по крайней мере в отношении одной переменной клетки расположены весьма упорядоченно. Речь идет о закономерном соответствии между распределением клеток в стриарной коре и положением их рецептивных полей на сетчатке, т.е. о том, что соседние клетки коры должны иметь рецептивные поля, расположенные близко друг к другу в поле зрения. Именно такая картина выявляется в экспериментах. У двух клеток, лежащих рядом в коре, рецептивные поля обычно даже перекрываются на большей части своей площади. Тем не менее эти поля не накладываются точно друг на друга. Если сдвигать микроэлектрод вдоль коры от клетки к клетке, то положения соответствующих рецептивных полей сдвигаются в направлении, которое можно предсказать, зная топографию отображения сетчатки в коре. Никто не усомнился бы в таком результате и 50 лет назад, имея данные о связях НКТ с корой и о случаях локальной слепоты после инсультов. Но как обстоит дело с остальными параметрами — глазодоминантностью, «сложностью», ориентацией адекватных стимулов и другими? Потребовалось несколько лет для того, чтобы научиться достаточно надежно стимулировать корковые клетки и регистрировать их ответы; в результате появилась возможность описывать реакции не только отдельных клеток, но и сравнительно больших групп нейронов. Началось с того, что нам случайно удалось одновременно записать ответы двух или нескольких клеток (пример такой записи был приведен на рис. 59). Записать ответ двух соседних клеток несложно. В тех экспериментах, где мы определяли предпочитаемый клеткой стимул, мы почти всегда использовали внеклеточное отведение, помещая кончик микроэлектрода рядом с клеткой; в этом случае регистрировались не изменения мембранного потенциала, а токи, связанные с импульсами. Часто при этом оказывалось, что мы регистрируем реакции не одной, а нескольких клеток одновременно — скажем, в том случае, когда кончик микроэлектрода останавливался на полпути между двумя телами нейронов. Импульсные разряды одиночных клеток при таком отведении почти идентичны, однако величина и форма импульсов зависят от расстояния и от взаимного расположения клеток, так что разряды, отводимые одновременно от двух клеток, обычно оказываются разными, и поэтому их можно легко различить. Выполняя такого рода отведения от двух клеток, мы смогли отчетливо увидеть, чем различаются соседние клетки и в чем они одинаковы. В одном из первых таких отведений были обнаружены две корковые клетки, отвечавшие на противоположные движения руки, которой махали перед животным. В этом случае две лежавшие бок о бок клетки давали разные (по существу, противоположные) реакции на движение стимула. Однако в других отношениях эти клетки почти наверное проявляли сходные свойства. Если бы я в 1956 году был уже подготовлен к изучению ориентационной избирательности, то я, весьма вероятно, обнаружил бы, что обе предпочитаемые ориентации близки к вертикальной, так как данные клетки хорошо отвечали на горизонтальные движения стимула. Тот факт, что обе они реагировали на возвратно-поступательное движение руки, означает, что положения рецептивных полей этих клеток приблизительно совпадали. Если бы я исследовал эти нейроны на глазодоминантность, то скорее всего этот параметр тоже оказался бы одинаковым. Уже в первых записях активности корковых нейронов нас поразило то, как часто две клетки, реакции которых можно регистрировать одновременно, одинаковы по глазодоминантности, сложности и, что самое удивительное, по оптимальной ориентации стимулов. Такие совпадения, вряд ли случайные, позволяют предположить, что клетки с одними и теми же свойствами объединены в группы. Возможность такой группировки весьма заинтересовала нас, и как только это предположение подтвердилось, мы стали выяснять, каковы размеры и форма этих групп.
Исследование коры
С помощью микроэлектродов можно исследовать только отдельные точки коры. Для того чтобы получить представление о трехмерной организации мозга, приходится медленно погружать электрод в глубину, время от времени останавливать его для записи активности какой-нибудь клетки (а возможно, — двух или трех клеток), отмечать по специальной шкале показания глубины, а затем повторять все сначала. Рано или поздно кончик микроэлектрода пройдет через весь корковый слой, и тогда электрод можно вынуть и снова ввести его в каком-нибудь другом месте. После окончания эксперимента делают срез, окрашивают его и исследуют под микроскопом с целью определить положение каждой из нервных клеток, активность которых регистрировалась. В одном эксперименте длительностью около 24 часов обычно удается сделать две-три проходки примерно по 4–5 миллиметров каждая. За одну проходку можно наблюдать ответы примерно 200 клеток. Микроэлектрод настолько тонок, что едва удается найти след от его прохождения под микроскопом, поэтому нет оснований думать, что при проходке микроэлектродом будет повреждено много клеток и это может повлиять на реакции близлежащих нейронов. Вначале нам было трудно находить на гистологическом срезе след от микроэлектрода, не говоря уже о том, чтобы определить положение кончика электрода, и это затрудняло оценку положения тех клеток, активность которых была записана. Эту трудность удалось преодолеть, когда было обнаружено, что пропускание через микроэлектрод слабого тока приводит к разрушению клеток в ближайшей окрестности кончика микроэлектрода, и на гистологических срезах эта зона разрушения хорошо видна. К счастью, при пропускании тока сам микроэлектрод не повреждается. Поэтому за одну проходку ток пропускают 3–4 раза, отмечая при этом глубину погружения микроэлектрода, а так как глубину отмечают и при регистрации активности клеток, то можно оценить и положение каждой из них. Разумеется, при воздействии тока погибает несколько клеток около кончика микроэлектрода, однако их не так много, чтобы могла быть нарушена работа более удаленных нейронов. Для того чтобы при этом не исказить ответы клеток, лежащих впереди на пути микроэлектрода, его кончик немного продвигают вперед, регистрируют активность клеток, а затем кончик отводят назад и тогда уже пропускают ток.
Вариации в сложности
Как и следовало ожидать, клетки во входном корковом слое — слое 4 — проявляют более простое поведение, чем клетки на выходе. Мы уже отмечали в этой главе, что у обезьяны клетки в слое 4Cβ, куда приходят волокна из четырех верхних (мелкоклеточных) слоев НКТ, по-видимому, не обладают избирательностью к ориентации стимула и ведут себя подобно клеткам, имеющим круглые рецептивные поля с центром и периферией. В слое 4Cα, имеющем входы от двух вентральных (крупноклеточных) слоев НКТ, некоторые клетки обладают круглыми полями с центром и периферией, а всем остальным, видимо, свойственны простые рецептивные поля и ориентационная чувствительность. Если перейти к следующему уровню — к слоям, лежащим выше и ниже слоя 4C, — то подавляющее большинство клеток окажутся сложными. Клетки, реагирующие на концы линий, в слоях 2 и 3 составляют около 20 процентов, а в других слоях встречаются редко. Таким образом, в целом обнаруживается явная корреляция между сложностью клеток и положением их в зрительном пути, которое можно оценить по числу синаптических переключений до данного места.
 Рис. 67. Примерная схема размещения клеток физиологически различного типа в разных слоях стриарной коры.
Рис. 67. Примерная схема размещения клеток физиологически различного типа в разных слоях стриарной коры.
В утверждении, что большинство клеток выше и ниже слоя 4 — сложные, упускаются из виду существенные различия между слоями: сложные клетки здесь далеко не одинаковы. Разумеется, все они имеют одно общее свойство, характерное для сложных клеток, — на движущуюся линию оптимальной ориентации они отвечают по всему рецептивному полю, независимо от конкретного положения стимула. Однако они различаются между собой другими свойствами. Можно выделить четыре подтипа клеток, которые содержатся в основном в разных слоях. Большинство сложных клеток в слоях 2 и 3 отвечает тем лучше, чем длиннее стимульная линия (в этом проявляется свойство суммации по длине). Однако реакция таких клеток становится слабее, когда длина стимула превышает некоторую критическую величину (если данная сложная клетка относится к нейронам, реагирующим на концы линий). Что касается клеток слоя 5, то для них короткие линии, занимающие по длине лишь небольшую долю рецептивного поля, почти так же эффективны, как и длинные; рецептивные поля этих клеток гораздо больше, чем у клеток в слоях 2 и 3. Напротив, клетки слоя 6 реагируют тем лучше, чем длиннее линия с оптимальной ориентацией (их ответ начинает ухудшаться только тогда, когда линия займет всю длину рецептивного поля, которая в несколько раз больше его ширины, т.е. рецептивное поле длинное и узкое). Можно сделать вывод, что аксоны, выходящие из слоев 5 и 6 и из слоев 2 и 3 и идущие к разным участкам мозга (к верхним бугоркам четверохолмия, к НКТ, к другим зрительным полям), должны служить для передачи разных видов зрительной информации. Подводя итоги, можно сказать, что при переходе от слоя к слою в поведении клеток выявляются более важные различия, чем, скажем, различия в оптимальной ориентации стимула или же в характере глазодоминантности. Наиболее заметные различия между клетками разных корковых слоев касаются сложности их реакций; это отражает тот простой анатомический факт, что одни слои расположены ближе к входу в кору, чем другие.
Колонки глазодоминантности
Группы клеток стриарной коры с различной глазодоминантностью были обнаружены раньше всего, так как они довольно крупные. Разработано много методов исследования таких группировок, поэтому они сейчас лучше всего изучены. Уже в самом начале исследований на коре обезьян стало очевидно, что всякий раз, когда микроэлектрод входит в кору перпендикулярно ее поверхности, он встречает одну за другой клетки, лучше реагирующие на стимуляцию одного и того же глаза (это показано на рис. 68). Если микроэлектрод вынуть и ввести его в другом месте в нескольких миллиметрах от предыдущего, то снова для всех встречающихся клеток доминирующим будет один глаз — тот же, что и раньше, или другой. В слое 4C, имеющем входы непосредственно от НКТ, доминирование одного глаза становится уже не относительным, а абсолютным, монопольным. В тех же слоях, которые расположены выше и ниже и, следовательно, находятся дальше в цепи синаптических переключений, более половины всех клеток могут возбуждаться и от недоминирующего глаза. Такие клетки мы называем бинокулярными.
 Рис. 68. Глазодоминантность остается неизменной при вертикальном продвижении микроэлектрода в стриарной коре. Если же электрод продвигается параллельно поверхности коры, то зоны с различной глазодоминантностью чередуются, причем каждая пара таких зон занимает около 1 мм.
Рис. 68. Глазодоминантность остается неизменной при вертикальном продвижении микроэлектрода в стриарной коре. Если же электрод продвигается параллельно поверхности коры, то зоны с различной глазодоминантностью чередуются, причем каждая пара таких зон занимает около 1 мм.
Если вместо того, чтобы перемещать электрод перпендикулярно поверхности коры, мы вводили его с наклоном, как можно ближе к линии, параллельной поверхности, то клетки с разной глазодоминантностью чередовались — доминирующим был то один глаз, то другой. Полный цикл этой смены соответствовал примерно одному миллиметру. Очевидно, если бы можно было видеть структуру коры сверху, она представлялась бы мозаикой, составленной из чередующихся зон с доминированием правого и левого глаза. Чем обусловлено такое чередование, стало ясно, когда был разработан новый метод окраски, позволяющий прослеживать ветвление отдельных аксонов, приходящих из НКТ, и распределение их окончаний в коре. Ветвление одного аксона таково, что тысячи его концевых синапсов образуют в слое 4C два или три скопления шириной 0,5 мм, разделенных промежутками около 0,5 мм, как показано на рис. 69. Поскольку клетки НКТ монокулярные, каждый отдельный аксон связан в конечном счете либо с правым, либо с левым глазом. Предположим, что аксон, изображенный на рис. 69 слева, относится к волокнам от левого глаза; в этом случае каждое входящее в тот же участок коры волокно, связанное с левым глазом, будет разветвляться в тех же скоплениях окончаний величиной 0,5 мм каждое. Промежутки между этими скоплениями размером тоже 0,5 мм будут заняты концевыми разветвлениями волокон от правого глаза. Такой особый тип распределения аксонов из НКТ в слое 4C сразу позволяет объяснить строгую монокулярность клеток в этом слое.
 Рис. 69. Каждый аксон из HKT сначала проходит в стриарной коре через нижние слои, последовательно ветвится и затем оканчивается в слое 4C. Здесь он образует скопления синаптических окончаний шириной 0,5 мм, отделенные друг от друга промежутками такой же ширины. Все волокна от одного глаза группируются в одних и тех же участках. В промежутках между этими участками группируются волокна от другого глаза. Область разветвлений одного волокна из крупноклеточного слоя HKT может составлять от 2 до 3 мм (окончания находятся в слое 4Cα). Волокно из мелкоклеточного слоя HKT ветвится в более ограниченной области (в слое 4Cβ), обычно занимающей лишь одну или две колонки.
Рис. 69. Каждый аксон из HKT сначала проходит в стриарной коре через нижние слои, последовательно ветвится и затем оканчивается в слое 4C. Здесь он образует скопления синаптических окончаний шириной 0,5 мм, отделенные друг от друга промежутками такой же ширины. Все волокна от одного глаза группируются в одних и тех же участках. В промежутках между этими участками группируются волокна от другого глаза. Область разветвлений одного волокна из крупноклеточного слоя HKT может составлять от 2 до 3 мм (окончания находятся в слое 4Cα). Волокно из мелкоклеточного слоя HKT ветвится в более ограниченной области (в слое 4Cβ), обычно занимающей лишь одну или две колонки.
Для того чтобы избирательно окрашивать одно и только одно волокно, требовался новый метод, и он был предложен в конце 1970-х годов. Этот метод основан на использовании аксонного транспорта — процесса, в результате которого различные материалы (белки или даже более крупные частицы) непрерывно переносятся в обоих направлениях внутри аксона. Некоторые из них транспортируются со скоростью нескольких сантиметров в час, другие — несколько миллиметров в сутки. Для того чтобы окрасить одиночный аксон, с помощью микропипетки вводят в него вещество, о котором известно, что оно переносится по аксону и окрашивает его, но не влияет на структуру клетки. В настоящее время чаще всего используют фермент пероксидазу из хрена. Этот фермент распространяется по аксону в обоих направлениях и катализирует химическую реакцию, продукт которой очень эффективно окрашивает клетку. Поскольку фермент служит катализатором, достаточно ничтожных его количеств, чтобы получить интенсивную окраску. Важно и то, что в самой нервной ткани фермента с подобными свойствами нет: это исключает возможность нежелательной окраски фона. Тот факт, что кора разбита на колонки глазодоминантности, проходящие от поверхности мозга вплоть до белого вещества, подтвердил морфологические данные о том, что группы клеток в слое 4C служат главными местами передачи зрительной информации в клеточные слои, лежащие выше и ниже этого слоя. Наличие некоторого числа горизонтальных и диагональных связей длиной около миллиметра, идущих по всем направлениям, должно приводить к некоторому размыванию четких зон доминирования правого или левого глаза в слоях, расположенных выше и ниже слоя 4C (рис. 70). Поэтому можно ожидать, что клетка, расположенная прямо над центром зоны доминирования левого глаза в слое 4, будет явно лучше отвечать на стимуляцию этого глаза (а возможно, и полностью контролироваться им), тогда как клетка, расположенная ближе к границе между участками разной глазодоминантности, может оказаться бинокулярной без какого-либо доминирования одного из глаз. Действительно, при горизонтальном передвижении электрода в верхнем слое коры или же в слое 5 или 6 отмечаются постепенные изменения глазодоминантности: сначала будут встречаться клетки, намного сильнее активируемые одним глазом, затем клетки, у которых эта асимметрия выражена слабее, потом клетки с равной зависимостью от обоих глаз, после чего начнет усиливаться влияние другого глаза. Такие плавные переходы резко контрастируют с той внезапной сменой глазодоминантности, которую мы наблюдаем при движении электрода вдоль слоя 4C (рис. 71).
 Рис. 70. Из-за наличия горизонтальных и диагональных связей выше слоя 4 происходит перекрывание колонок глазодоминантности и границы их размываются.
Рис. 70. Из-за наличия горизонтальных и диагональных связей выше слоя 4 происходит перекрывание колонок глазодоминантности и границы их размываются.
 Рис. 71. В отличие от четких границ между колонками глазодоминантности в слое 4 коры, границы колонок в верхних (2 и 3) и нижних (5 и 6) слоях коры размыты. Стрелками показано направление продвижения микроэлектрода в слое 4 (вверху слева) и в слое 2 или 3 (вверху справа). Внизу представлена глазодоминантность клеток, ответы которых регистрировались в одной проходке микроэлектрода. В слое 4 видны резкие переходы от группы 1 (влияет только контралатеральный глаз) к группе 7 (влияет только ипсилатеральный глаз). В других слоях встречается много бинокулярных клеток, а глазодоминантность периодически изменяется, проходя через промежуточные уровни (цифры 1, 4 и 7 означают категории клеток по глазодоминантности).
Рис. 71. В отличие от четких границ между колонками глазодоминантности в слое 4 коры, границы колонок в верхних (2 и 3) и нижних (5 и 6) слоях коры размыты. Стрелками показано направление продвижения микроэлектрода в слое 4 (вверху слева) и в слое 2 или 3 (вверху справа). Внизу представлена глазодоминантность клеток, ответы которых регистрировались в одной проходке микроэлектрода. В слое 4 видны резкие переходы от группы 1 (влияет только контралатеральный глаз) к группе 7 (влияет только ипсилатеральный глаз). В других слоях встречается много бинокулярных клеток, а глазодоминантность периодически изменяется, проходя через промежуточные уровни (цифры 1, 4 и 7 означают категории клеток по глазодоминантности).
Если смотреть «сбоку», две совокупности клеток в слое 4C выглядят как чередующиеся участки. Однако нам хотелось узнать, как будет выглядеть распределение этих участков при взгляде на поверхность коры сверху. Допустим, что они отмечены на поверхности коры черным или белым цветом. С точки зрения топологии можно распределить эти участки несколькими разными способами: в шахматном порядке, в виде чередующихся черных и белых полос, в виде черных пятен на белом фоне или использовать любые комбинации этих паттернов. На рис. 72 показаны три возможных варианта такого распределения. Решать поставленную задачу с помощью одних только микроэлектродов означает использовать одномерные средства для расшифровки неизвестной трехмерной структуры. Это задача безнадежная — все равно что пытаться подстричь траву на лужайке с помощью ножниц для ногтей. Столкнувшись с такими трудностями, многие предпочли бы переключиться на совершенно иной вид деятельности — заняться, скажем, сельским хозяйством или юриспруденцией. (В начале 1960-х годов, когда Торстен Визел и я были более терпеливыми и решительными, чем сейчас, мы действительно попытались выяснить топографию глазодоминантности с помощью микроэлектродов, и нам удалось даже получить кое-какие результаты. Образно выражаясь, я в те дни действительно пытался подстричь травяной газон ножницами, так как мы не могли позволить себе купить газонокосилку. В то время мы были беднее, чем нынешние аспиранты, однако были, вероятно, более настойчивыми.)
 Рис. 72. Здесь показаны три возможных способа разбиения поверхности на два вида областей — разбиение в виде шахматной доски, в виде полос и в виде островков. В данном случае речь идет о поверхности коры и зонах доминирования правого или левого глаза.
Рис. 72. Здесь показаны три возможных способа разбиения поверхности на два вида областей — разбиение в виде шахматной доски, в виде полос и в виде островков. В данном случае речь идет о поверхности коры и зонах доминирования правого или левого глаза.
К счастью, в последние десять лет были придуманы очень эффективные методы нейроморфологических исследований, и сейчас поставленная нами задача уже решена независимо несколькими способами; два из них описаны ниже. Первый способ опять-таки основан на использовании аксонного транспорта. Берут небольшое количество органического вещества (например, аминокислоты́), меченного радиоактивным изотопом (например, 14C), и инъецируют в один глаз обезьяны, скажем в левый. Вещество захватывается клетками глаза, в том числе и ганглиозными клетками сетчатки. Помеченные молекулы, по-видимому уже включенные в состав белков, переносятся по аксонам ганглиозных клеток и их окончаниям в НКТ. Здесь они накапливаются в слоях, связанных с левым глазом. Для переноса метки требуется несколько дней. Затем из ткани приготовляют тонкие срезы, накладывают на них фотоэмульсию и оставляют на несколько дней в темноте. В результате получается радиоавтограф вроде показанного на рис. 73, где можно увидеть в каждом из двух НКТ по три слоя, связанных с левым глазом. Черные области, где образовались зерна, в одном НКТ расположены так, как светлые области в другом.
 Рис. 73. На этих срезах наружных коленчатых тел — левого и правого — можно видеть концентрацию радиоактивной метки в трех слоях, связанных с левым глазом. За неделю до получения срезов в левый глаз был введен пролин, меченный тритием. Те слои, в которых накопилась метка, выглядят на радиоавтографе темными и более толстыми.
Рис. 73. На этих срезах наружных коленчатых тел — левого и правого — можно видеть концентрацию радиоактивной метки в трех слоях, связанных с левым глазом. За неделю до получения срезов в левый глаз был введен пролин, меченный тритием. Те слои, в которых накопилась метка, выглядят на радиоавтографе темными и более толстыми.
Для того чтобы наблюдать такую картину организации НКТ, нужно ввести в глаз не так много радиоактивного вещества. Если же при инъекции ввести его в достаточно большом количестве, то концентрация его в слоях НКТ станет настолько высокой, что часть радиоактивной метки перейдет из окончаний волокон зрительного нерва в клетки самих НКТ (в пределах меченых слоев) и будет транспортироваться по их аксонам в стриарную кору. В результате метка начнет накапливаться в окончаниях в слое 4C, образуя регулярный узор, соответствующий тому глазу, в который была сделана инъекция. Когда будет готов радиоавтограф (для этого нужно несколько месяцев, так как концентрация меченого вещества, дошедшего до коры, очень низка), мы действительно увидим на поперечном срезе коры места накопления метки в слое 4C (рис. 74). Если же сделать срез коры параллельно ее поверхности (сначала распрямив кору или же разрезав ее на части и скрепив затем вместе последовательные срезы), то мы, наконец, сможем увидеть распределение колонок так, как если бы смотрели на поверхность коры сверху. Четко выраженный набор параллельных полосок можно видеть как на одиночном срезе (рис. 75, вверху), так и на реконструированном изображении (рис. 75, внизу). На всех этих радиоавтографах коры меченые участки, т.е. проекции левого глаза, выглядят светлыми, а немеченые промежутки между ними — проекции правого глаза — темными. Поскольку из слоя 4 пути к верхним и нижним слоям идут главным образом в вертикальном направлении, прямо вверх и вниз, то в трехмерном пространстве зоны глазодоминантности образуют систему чередующихся «ломтиков», соответствующих правому и левому глазу (рис. 76).
 Рис. 74. На этом радиоавтографе среза стриарной коры светлые участки соответствуют тем местам в слое 4, где накопилось много радиоактивного вещества после инъекции его в левый глаз животного. Эти участки разделены темными зонами, не содержащими радиоактивной метки, — зонами доминирования правого глаза.
Рис. 74. На этом радиоавтографе среза стриарной коры светлые участки соответствуют тем местам в слое 4, где накопилось много радиоактивного вещества после инъекции его в левый глаз животного. Эти участки разделены темными зонами, не содержащими радиоактивной метки, — зонами доминирования правого глаза.
 Рис. 75. Вверху: срез куполообразного участка коры, проходящий параллельно поверхности мозга. Слой 4, пересекаясь со срезом, образует кольцо. Внизу: результат объединения нескольких таких колец, полученных на серии срезов (чем глубже сделан срез, тем больше диаметр кольца). (Картина получается не очень четкой, так как трудно сфотографировать все срезы в одинаковых условиях, особенно если учесть то, что автор — всего лишь фотолюбитель).
Рис. 75. Вверху: срез куполообразного участка коры, проходящий параллельно поверхности мозга. Слой 4, пересекаясь со срезом, образует кольцо. Внизу: результат объединения нескольких таких колец, полученных на серии срезов (чем глубже сделан срез, тем больше диаметр кольца). (Картина получается не очень четкой, так как трудно сфотографировать все срезы в одинаковых условиях, особенно если учесть то, что автор — всего лишь фотолюбитель).
 Рис. 76. В объемном представлении колонки глазодоминантности больше похожи не на греческие колонны, а на ломтики нарезанного хлеба, расположенные перпендикулярно поверхности.
Рис. 76. В объемном представлении колонки глазодоминантности больше похожи не на греческие колонны, а на ломтики нарезанного хлеба, расположенные перпендикулярно поверхности.
Используя другой способ, С. Ле-Вэй успешно осуществил реконструкцию вида всей стриарной коры в затылочной доле. Участок коры, лежащий на поверхности мозга, показан на рис. 77. Распределение полос оказалось наиболее регулярным и четким на некотором расстоянии от корковой проекции центральной ямки. Вблизи этой проекции узор по неизвестным причинам наиболее сложен — он отличается регулярностью, но содержит много петель и завитков, поэтому он здесь мало похож на тот правильный узор, напоминающий рисунок обоев, который характерен для областей коры, более удаленных от проекции центральной ямки. Ширина полосок везде постоянна и равна примерно 0,5 мм. В процентном отношении корковые представительства правого и левого глаза примерно равны, пока речь идет о проекции центральной области сетчатки диаметром около 40°. Ле-Вэй и Д. ван Эссен обнаружили, что за пределами этой области из-за уменьшения вклада ипсилатерального глаза соответствующие ему полоски сужаются до 0,25 мм. Если же говорить о проекции зоны, удаленной от центральной ямки на 70–80°, то здесь, разумеется, представлен только ипсилатеральный глаз. Это естественно, так как поле зрения правого глаза простирается вправо дальше, чем влево.
 Рис. 77. Здесь показан результат реконструкции зон глазодоминантности по Ле-Вэю (часть поля 17 правого полушария).
Рис. 77. Здесь показан результат реконструкции зон глазодоминантности по Ле-Вэю (часть поля 17 правого полушария).
Второй метод изучения колонок глазодоминантности позволяет выявлять их во всей толще коры, а не только в слое 4. Это метод с использованием меченой 2-дезоксиглюкозы, изобретенный в 1976 году Л. Соколоффом в Национальных институтах здоровья в Бетезде. Он тоже основан на способности радиоактивных веществ засвечивать фотопленку. В основе его лежит тот факт, что нейроны, как и большинство других клеток организма, поглощают глюкозу в качестве источника энергии, и чем интенсивнее им приходится работать, тем больше глюкозы они потребляют. Поэтому можно было бы представить себе следующую процедуру. Животному впрыскивают радиоактивную глюкозу, а затем стимулируют один глаз, например правый, в течение нескольких минут предъявляя какую-либо фигуру (время стимуляции должно быть достаточным для того, чтобы возбужденные клетки в мозгу поглотили введенную глюкозу). После этого мозг извлекают, делают срезы и покрывают их фотоэмульсией, а затем после надлежащей экспозиции получают, как и в предыдущем случае, радиоавтограф. Однако на самом деле такая схема не годится, так как глюкоза, поглощенная клетками, расщепляется с образованием продуктов, которые быстро переходят обратно в кровь. Для того чтобы предотвратить эту утечку, Соколофф предложил остроумный прием — использовать при инъекции дезоксиглюкозу, которая по химической структуре очень близка к обычной глюкозе. «Обманутые» клетки поглощают ее и даже пытаются расщеплять; однако этот процесс прерывается после первого же шага; дезоксиглюкоза превращается в 2-дезоксиглюкозо-6-фосфат, который далее не может быть расщеплен. К счастью, это вещество нерастворимо в липидах клеточной мембраны и поэтому не может выйти из клетки. Оно накапливается в клетке в таком количестве, что уже может быть обнаружено с помощью радиоавтографии. Рассматривая результат этой процедуры — радиоавтограф, можно получить представление о том, какие участки мозга были наиболее активны в период стимуляции и накопили больше всего этой «фальшивой глюкозы». Если бы в ходе опыта животное делало движения лапой, то на радиоавтографе выявился бы также соответствующий участок моторной зоны коры. То, что мы видим после стимуляции правого глаза, — это та часть вещества коры, которая была в наибольшей степени возбуждена данным стимулом, а именно набор колонок глазодоминантности для правого глаза. Примеры получаемых результатов представлены на рис. 78.
 Рис. 78. Результаты двух экспериментов с радиоактивной дезоксиглюкозой. Вверху: срез, проходящий через затылочные доли обоих полушарий животного, воспринимавшего зрительные стимулы обоими глазами после внутривенного введения метки. Внизу: после инъекции животное воспринимало стимул лишь одним глазом (другой был закрыт). В этом случае в коре хорошо заметны колонки глазодоминантности. (Эксперименты C. Kennedy, M. N. Des Rosiers, O. Sakurada, M. Shinohara, M. Reivich, T. W. Tehle, L. Sokoloff.)
Рис. 78. Результаты двух экспериментов с радиоактивной дезоксиглюкозой. Вверху: срез, проходящий через затылочные доли обоих полушарий животного, воспринимавшего зрительные стимулы обоими глазами после внутривенного введения метки. Внизу: после инъекции животное воспринимало стимул лишь одним глазом (другой был закрыт). В этом случае в коре хорошо заметны колонки глазодоминантности. (Эксперименты C. Kennedy, M. N. Des Rosiers, O. Sakurada, M. Shinohara, M. Reivich, T. W. Tehle, L. Sokoloff.)
Идею того же метода исследования удачно развил Р. Тутелл из лаборатории Рассела де Валуа в Беркли, взяв в качестве стимула для животного, смотревшего на экран одним глазом, большую фигуру в виде концентрических окружностей и радиальных линий (рис. 79, вверху). Получившаяся в результате картина корковой проекции тоже содержала круги и радиусы, но только в искаженном виде, что связано с неодинаковым увеличением разных участков сетчатки в их проекции на стриарную кору (с этим же связано и различие в остроте зрения на периферии сетчатки и в центральной ямке). Кроме того, каждая из окружностей и каждый из радиусов разбиты на множество мелких участков, соответствующих колонкам глазодоминантности. При одновременной стимуляции обоих глаз полосы были бы непрерывными. Редко так бывает, что в одном эксперименте удается столь четко продемонстрировать сразу несколько важных фактов.
 Рис. 79. В этом эксперименте, проведенном Р. Тутеллом, в центре поля зрения наркотизированного макака в течение 45 мин предъявляли стимул, напоминающий мишень с несколькими радиальными линиями. Предварительно животному была сделана инъекция радиоактивной 2-дезоксиглюкозы. Один глаз был закрыт. В нижней части рисунка показано распределение метки в стриарной коре левого полушария мозга. На этом радиоавтографе представлен срез коры, параллельный ее поверхности. Перед тем как сделать срез, корковую ткань растянули и заморозили. Полукруглые линии стимула отображаются в коре как почти вертикальные линии, а радиальные линии правой части зрительного поля — в виде горизонтальных линий. «Пунктирный» характер каждой линии на радиоавтографе обусловлен тем, что в опыте стимулировался только один глаз и, значит, возбуждались только соответствующие колонки глазодоминантности.
Рис. 79. В этом эксперименте, проведенном Р. Тутеллом, в центре поля зрения наркотизированного макака в течение 45 мин предъявляли стимул, напоминающий мишень с несколькими радиальными линиями. Предварительно животному была сделана инъекция радиоактивной 2-дезоксиглюкозы. Один глаз был закрыт. В нижней части рисунка показано распределение метки в стриарной коре левого полушария мозга. На этом радиоавтографе представлен срез коры, параллельный ее поверхности. Перед тем как сделать срез, корковую ткань растянули и заморозили. Полукруглые линии стимула отображаются в коре как почти вертикальные линии, а радиальные линии правой части зрительного поля — в виде горизонтальных линий. «Пунктирный» характер каждой линии на радиоавтографе обусловлен тем, что в опыте стимулировался только один глаз и, значит, возбуждались только соответствующие колонки глазодоминантности.
Колонки глазодоминантности есть у кошек, некоторых низших обезьян, шимпанзе и человека. У грызунов и тупайи их нет. У саймири (род обезьян Нового Света) некоторые указания на возможное существование таких колонок получены в физиологических экспериментах, но современные методы морфологического анализа не выявляют их. В настоящее время нам неизвестна роль этого сложного распределения сигналов, приходящих от разных глаз; возможно, что оно имеет какое-то отношение к стереоскопическому зрению. Подразделение на участки с функционально специализированными клетками было обнаружено не только в стриарной коре. Впервые такие участки описал в середине 1960-х годов В. Маунткасл в соматосенсорной коре. Это явилось наиболее важным открытием с того времени, когда были получены первые сведения о локализации функций мозга. Соматосенсорная область коры имеет такое же отношение к осязанию и проприоцепции, как стриарная кора — к зрению. Маунткасл показал, что эта область подразделяется на вертикально ориентированные зоны, внутри которых клетки чувствительны к прикосновению, и зоны, в которых клетки отвечают на сгибание в суставах или на приложение значительного давления к конечности. Так же как и в случае колонок глазодоминантности, ширина этих зон составляет примерно 0,5 мм. Однако еще не ясно, образуют ли эти зоны полосы, шахматный рисунок или же отдельные островки на общем фоне. Маунткасл назвал их колонками, и можно думать, что он мысленно представлял себе некую сотовидную структуру. Теперь мы уже знаем, что для зрительной коры более подходящим был бы термин пластина или слэб (slab). Однако введенную терминологию очень трудно изменить, поэтому лучше всего, наверное, сохранить прежний термин, несмотря на его недостатки. Сегодня мы говорим о колончатой организации, когда некоторое свойство клеток остается постоянным во всей толще коры от ее поверхности до белого вещества, но изменяется в направлениях, параллельных поверхности коры. По причинам, которые будут ясны из следующей главы, мы обычно не применяем этот термин, когда речь идет о топографическом отображении, т.е. о проекции расположения рецептивных полей на сетчатке или на поверхности тела.
Ориентационные колонки
Регистрируя реакции нейронов стриарной коры, мы уже в самом начале заметили, что всякий раз, когда одновременно отводится активность двух клеток, эти клетки оказываются сходными не только по глазодоминантности, но и по оптимальной ориентации стимула. Возникает вопрос: однотипны ли соседние клетки и по всем другим свойствам? Ответ будет отрицательным. Как я уже упоминал, положения рецептивных полей в большинстве случаев не вполне совпадают, хотя поля обычно перекрываются; дирекциональная чувствительность часто бывает противоположной, или же у одной клетки она может быть хорошо выражена, а у другой ее может не быть вовсе. В слоях 2 и 3, где встречаются клетки, реагирующие на концы линий, одна клетка может не проявлять совсем этого свойства, а соседняя — обладать им в полной мере. С другой стороны, две соседние клетки очень редко обнаруживают явное различие в оптимальной ориентации стимула или противоположную глазодоминантность. Предпочитаемая ориентация, так же как и глазодоминантность, остается неизменной при прохождении электрода вертикально сквозь всю толщу коры. Как уже говорилось, в слое 4Cβ клетки вообще не обладают избирательностью к ориентации стимула; но как только мы доходим до слоя 5, у клеток выявляется сильно выраженная настройка на определенную ориентацию, причем оптимальная ориентация здесь та же, что была выше слоя 4. Если теперь вынуть электрод и ввести его в каком-нибудь другом месте, то общая картина останется прежней, только ориентация скорее всего будет уже другой. Таким образом, кора разбита на узкие участки с постоянной предпочтительной ориентацией, которые идут от поверхности коры до белого вещества, но прерываются в слое 4, где клетки не обладают ориентационной избирательностью. Если же, наоборот, вводить электрод параллельно поверхности коры, то наблюдается удивительно закономерное изменение предпочитаемой ориентации — каждый раз, когда электрод перемещается на 0,05 мм (50 мкм), ориентация сдвигается в среднем на 10° по часовой стрелке или против часовой стрелки. Поэтому при продвижении электрода на 1 мм она обычно меняется на противоположную. Величины 50 мкм и 10° близки к пределу доступной ныне точности измерений, так что нельзя сказать определенно, меняется ли ориентация при смещении электрода непрерывно или же сдвигается скачками. На рис. 80 и 81 представлена часть типичного эксперимента, в котором электрод продвигался в поле 17 в направлении, близком к горизонтальному. В этом опыте точки фиксации двух глаз на экране не вполне совпадали (из-за наркоза и введения вещества, расслабляющего мышцы) — расстояние между ними составляло около 2°. Цветные круги на рис. 86 примерно соответствуют размерам рецептивных полей (около 1° в диаметре), расположенных в 4° ниже и слева от центральных ямок (регистрировались ответы клеток правого полушария). Первая клетка, отмеченная номером 96, была бинокулярной, однако для следующей, 114, явно доминирующим оказался правый глаз. Затем идут клетки под номерами от 111 до 118 с доминированием левого глаза. Легко заметить, как регулярно изменяется ориентация в последовательности клеток, в данном случае в направлении против часовой стрелки. Если построить график зависимости ориентации от продвижения электрода (рис. 87), то все точки лягут на линию, близкую к прямой. Переход от одного глаза к другому не сопровождался никакими резкими изменениями ни в направлении сдвига ориентации, ни в наклоне линии на графике. Согласно нашей интерпретации, это означает, что между двумя системами группировки — по глазодоминантности и по ориентации стимула — нет тесной связи. Это выглядит так, как если бы кора размечалась двумя совершенно независимыми способами.
 Рис. 80. Если ввести микроэлектрод в поле 17 коры макака очень наклонно, выявляется весьма регулярное изменение ориентационной избирательности (в данном случае исследованы 23 соседние клетки).
Рис. 80. Если ввести микроэлектрод в поле 17 коры макака очень наклонно, выявляется весьма регулярное изменение ориентационной избирательности (в данном случае исследованы 23 соседние клетки).
 Рис. 81. Графическое представление результатов, отображенных на рис. 80. Это график зависимости оптимальной ориентации стимула (в угловых градусах) от расстояния, пройденного микроэлектродом. (Поскольку электрод вводился почти параллельно поверхности коры, длина проходки почти такая же, как соответствующее расстояние на поверхности коры). В этом эксперименте полный поворот ориентации стимула на 180° происходил на пути в 0,7 мм.
Рис. 81. Графическое представление результатов, отображенных на рис. 80. Это график зависимости оптимальной ориентации стимула (в угловых градусах) от расстояния, пройденного микроэлектродом. (Поскольку электрод вводился почти параллельно поверхности коры, длина проходки почти такая же, как соответствующее расстояние на поверхности коры). В этом эксперименте полный поворот ориентации стимула на 180° происходил на пути в 0,7 мм.
Когда электрод продвигается параллельно поверхности коры, оптимальная ориентация стимула может изменяться либо по часовой стрелке, либо в обратном направлении. В большинстве случаев при достаточно длинном пути электрода направление сдвига ориентации рано или поздно меняется на обратное. Момент такой смены, или реверсии, непредсказуем, но обычно она происходит с интервалами в несколько миллиметров. На рис. 82 приведен пример таких последовательных смен направления.
 Рис. 82. Результаты еще одного эксперимента, представленные в виде графика зависимости оптимальной ориентации стимула от расстояния, пройденного электродом. Видны три точки реверсии, в интервалах между которыми сохраняется прямая пропорциональность.
Рис. 82. Результаты еще одного эксперимента, представленные в виде графика зависимости оптимальной ориентации стимула от расстояния, пройденного электродом. Видны три точки реверсии, в интервалах между которыми сохраняется прямая пропорциональность.
Наконец, в некоторых экспериментах мы обнаружили еще одну особенность, которую назвали разрывом. Как раз тогда, когда нас начинала гипнотизировать монотонная регулярность постепенного изменения ориентации в одном и том же направлении, этот ход событий вдруг — в редких случаях — прерывался, и происходил сдвиг ориентации сразу на 45–90°. После этого опять шла прежняя регулярная последовательность, только направление ее часто менялось на обратное. На рис. 83 приведен пример таких разрывов, находящихся друг от друга на расстоянии в несколько десятых миллиметра.
 Рис. 83. Случай появления двух разрывов, т.е. точек, в которых оптимальная ориентация внезапно изменяется. До и после этих точек наблюдается плавное изменение оптимальной ориентации.
Рис. 83. Случай появления двух разрывов, т.е. точек, в которых оптимальная ориентация внезапно изменяется. До и после этих точек наблюдается плавное изменение оптимальной ориентации.
Как выглядели бы участки постоянной ориентации при взгляде на кору сверху? Ответить на этот вопрос оказалось гораздо труднее, чем на аналогичный вопрос о колонках глазодоминантности. Вплоть до самого последнего времени мы не имели прямой возможности «видеть» ориентационные группировки и могли лишь попытаться логически вывести их форму из результатов микроэлектродных исследований вроде описанных выше. Наличие как реверсий, так и разрывов наводит на мысль о сложности интересующей нас конфигурации. С другой стороны, линейная регулярность, которую мы наблюдали часто миллиметр за миллиметром вдоль коры, может означать периодичность, по крайней мере в пределах небольших зон коры; тогда реверсии и разрывы могли бы указывать на то, что эта периодичность прерывается через каждые несколько миллиметров. В тех областях, где периодичность сохраняется, можно с определенной вероятностью реконструировать искомую конфигурацию. Предположим, эта конфигурация такова, что в пределах данной области при продвижении электрода параллельно поверхности коры мы наблюдаем периодичность без реверсий и разрывов, т.е. везде получаем такой график, как на рис. 81. Если бы мы имели достаточное число таких графиков, то смогли бы реконструировать трехмерную структуру (рис. 84); здесь вертикальная ось (z) отображает ориентацию, а горизонтальные оси (x и y) — расстояния в плоскости коры. В этом случае ориентации отображались бы на некоторой поверхности; например, если график — прямая линия, то эта поверхность имела бы вид наклонной плоскости, что показано на рис. 84, а в других случаях это была бы криволинейная поверхность. На таких трехмерных реконструкциях искомые поверхности пересекались бы горизонтальными плоскостями (плоскостью x — y или плоскостями, параллельными ей) по некоторым линиям, соответствующим постоянной ориентации (можно назвать эти линии изоориентационными), что аналогично изогипсам на географических картах. Неровности рельефа — холмы, впадины, горы — на такой трехмерной реконструкции дадут в некоторых участках реверсии на графиках зависимости ориентации от расстояния, а резкие перепады высот в виде отвесных скал приведут к разрывам. Основной вывод из этих рассуждений заключается в том, что наличие зон регулярности означает возможность построить карту с изолиниями, откуда следует, что области постоянной ориентации, если рассматривать их сверху, должны иметь вид полос. Поскольку при вертикальной проходке коры предпочитаемые ориентации остаются неизменными, в объемном представлении эти области должны иметь вид пластин. А поскольку изоориентационные линии могут быть криволинейными, эти пластины не обязательно будут телами правильной формы наподобие ровно нарезанных ломтей хлеба. Многое из сказанного выше было продемонстрировано в прямых экспериментах, в которых производились две или три проходки в параллельных направлениях на расстоянии не более миллиметра друг от друга. Результатом этих экспериментов была реконструкция объемной картины по крайней мере в пределах небольшой исследованной области.
 Рис. 84. Трехмерное представление, где поверхность коры расположена в плоскости x-y, а оптимальная ориентация стимула откладывается по вертикальной оси z. Если при всех направлениях движения электрода получается линейная зависимость между ориентацией и расстоянием, то получающаяся в результате поверхность будет плоскостью и будет пересекаться с плоскостью x-y (если поверхность небудет плоской, она все равно будет пересекаться с этой плоскостью). В этом случае при проведении плоскостей, параллельных плоскости x-y, будут получаться изолинии. (Это утверждение выглядит на первый взгляд более сложным, чем оно есть на самом деле! Так же можно рассуждать и в том случае, если плоскость x-y будет поверхностью Огненной Земли, а по оси z будет откладываться высота, среднее количество осадков в январе или температура.)
Рис. 84. Трехмерное представление, где поверхность коры расположена в плоскости x-y, а оптимальная ориентация стимула откладывается по вертикальной оси z. Если при всех направлениях движения электрода получается линейная зависимость между ориентацией и расстоянием, то получающаяся в результате поверхность будет плоскостью и будет пересекаться с плоскостью x-y (если поверхность небудет плоской, она все равно будет пересекаться с этой плоскостью). В этом случае при проведении плоскостей, параллельных плоскости x-y, будут получаться изолинии. (Это утверждение выглядит на первый взгляд более сложным, чем оно есть на самом деле! Так же можно рассуждать и в том случае, если плоскость x-y будет поверхностью Огненной Земли, а по оси z будет откладываться высота, среднее количество осадков в январе или температура.)
Если наши рассуждения правильны, то при выполнении множества произвольных проходок некоторые из них случайно будут совпадать с направлением ориентационных изолиний, и тогда ориентация должна оставаться неизменной. Такое действительно случается, но не очень часто. Этого и следовало ожидать, так как тригонометрия подсказывает, что даже небольшое отклонение направления проходки от изолинии должно приводить к довольно большим изменениям в ориентации; поэтому лишь в очень немногих случаях графики зависимости ориентации от глубины введения электрода могут оказаться горизонтальными линиями. Число градаций ориентации, представленных в одном квадратном миллиметре коры, можно определить по наиболее крутым графикам, получаемым в эксперименте. Эта цифра составляет примерно 400 градусов на 1 мм, откуда следует, что полный набор ориентаций в пределах 180 градусов может содержаться на участке длиной около 0,5 мм. Эту величину нужно будет вспомнить, когда мы вернемся к вопросу о топографии коры и ее удивительной однородности. Однако, забегая вперед, хочу отметить, что толщина пары колонок глазодоминантности составляет примерно 0,4 + 0,4 мм, т.е. близка к 1 мм. Это вдвое больше толщины полного набора ориентационных пластин, однако порядок величины тот же. Вскоре, однако, мы воспользовались методом картирования с помощью дезоксиглюкозы, который позволил непосредственно изучать геометрию ориентационных колонок. Для этого в качестве стимула мы взяли просто рисунок из параллельных полос с неизменной ориентацией, скажем вертикальной, который предъявляли на протяжении всего эксперимента. Полученная при этом картина (рис. 85) оказалась гораздо более сложной, чем картина колонок глазодоминантности. Однако и здесь отчетливо была видна регулярность распределения с периодом 1 мм или чуть меньше (расстояние от середины одной темной полосы до середины следующей). Это согласуется с электрофизиологическими данными (с тем расстоянием, на которое нужно передвинуть электрод от клеток с определенной ориентацией, скажем вертикальной, чтобы через все промежуточные варианты снова дойти до вертикальной ориентации). В некоторых местах мы находили структуру в виде правильных полос на площади в несколько квадратных миллиметров. Мы задавались вопросом: нет ли какой-то связи между конфигурациями ориентационных полос и полос глазодоминантности — может быть, например, они идут параллельно или же пересекаются под прямым углом? В том же самом эксперименте нам удалось выявить глазодоминантные колонки, после того как в глаз животного была введена радиоактивная аминокислота, а затем полученные срезы ткани анализировались с помощью двух методик (результат показан на рис. 86). Мы не смогли обнаружить никакой явной корреляции. Сравнивая сложную картину ориентационных колонок с гораздо более простым распределением колонок глазодоминантности, очень трудно было решить, какая могла бы быть связь между ними.
 Рис. 85. После инъекции дезоксиглюкозы наркотизированной обезьяне предъявлялся стимул в виде медленно движущихся черных и белых полос. На полученном радиоавтографе можно видеть периодическое распределение метки, в частности в слоях 5 и 6 (обширная область слева от центра). Круглое пятно, расположенное еще дальше влево, обусловлено равномерным распределением метки и соответствует слою 4Cβ; этого и следовало ожидать, поскольку клетки в этом слое нечувствительны к ориентации.
Рис. 85. После инъекции дезоксиглюкозы наркотизированной обезьяне предъявлялся стимул в виде медленно движущихся черных и белых полос. На полученном радиоавтографе можно видеть периодическое распределение метки, в частности в слоях 5 и 6 (обширная область слева от центра). Круглое пятно, расположенное еще дальше влево, обусловлено равномерным распределением метки и соответствует слою 4Cβ; этого и следовало ожидать, поскольку клетки в этом слое нечувствительны к ориентации.
 Рис. 86. Тому же животному, что и в предыдущем случае, в глаз неделей раньше была введена радиоактивная аминокислота (пролин). После промывания среза в воде с целью растворить 2-дезоксиглюкозу был получен радиоавтограф. Радиоактивная метка выявляет здесь колонки глазодоминантности. Эти колонки не имеют никакой видимой связи с ориентационными колонками.
Рис. 86. Тому же животному, что и в предыдущем случае, в глаз неделей раньше была введена радиоактивная аминокислота (пролин). После промывания среза в воде с целью растворить 2-дезоксиглюкозу был получен радиоавтограф. Радиоактивная метка выявляет здесь колонки глазодоминантности. Эти колонки не имеют никакой видимой связи с ориентационными колонками.
У метода с применением дезоксиглюкозы есть один серьезный недостаток: никогда нет полной уверенности в том, что полученная картина распределения действительно связана с интересующим нас свойством стимула. Например, когда в качестве стимула используется рисунок из черных и белых вертикальных полос, как можно убедиться, что получившееся распределение связано именно с ориентацией этих полос, т.е. что в темных областях находятся клетки, отвечающие на вертикальные линии, а в светлых — клетки, которые на них не реагируют? Ведь вместо этого решающей особенностью данного стимула могло бы быть то, что он черно-белый, а не цветной, или же то, что он состоит из широких, а не узких полос, или, наконец, то, что экран расположен именно на таком, а не ином расстоянии от животного. Одно из косвенных подтверждений того, что действие дезоксиглюкозы затрагивает механизмы настройки на ориентацию, — это отсутствие каких-либо пятен или полос в слое 4C, где клетки не обладают избирательностью к ориентации. Другим подтверждением служит результат, полученный в исследованиях М. Страйкера из Калифорнийского университета в Сан-Франциско. Страйкер делал протяженные проходки микроэлектродом в стриарной коре кошек параллельно ее поверхности и каждый раз, когда встречались клетки с определенной оптимальной ориентацией, производил локальное разрушение ткани; затем после инъекции радиоактивной дезоксиглюкозы включался стимул с полосками одинаковой ориентации. В этих экспериментах была обнаружена явная корреляция между получаемым распределением метки на гистологическом срезе и ориентацией стимула. Наиболее четко удалось недавно выявить ориентационные колонки с помощью красителей, чувствительных к электрическому напряжению, которые в течение многих лет разрабатывал Л. Коэн в Йеле. Г. Блэсдел из Питтсбургского университета применил их при исследовании коры головного мозга. При использовании этого метода краситель, чувствительный к напряжению и окрашивающий клеточные мембраны, наносят на поверхность коры наркотизированного животного, и нервные клетки поглощают его. Когда в эксперименте животному предъявляют стимул, все реагирующие на него клетки изменяют свой цвет. Если в области, близкой к поверхности коры, таких клеток окажется достаточно много, эти изменения цвета можно уловить с помощью современных методов обработки телевизионных изображений. Хотя изменения цвета едва заметны, с помощью чувствительных телекамер и машинных методов фильтрации шумов их можно обнаружить. Блэсдел использовал в своих опытах стимуляцию полосами определенной ориентации, после чего фотографировал полученное распределение активности клеток на участке площадью в несколько квадратных сантиметров. Затем та же процедура повторялась при многих других ориентациях стимула. После этого каждой ориентации был поставлен в соответствие свой цвет — красный для вертикальной, оранжевый для положения часовой стрелки в 1 час и т.д.; потом все изображения были наложены друг на друга. Поскольку изоориентационная линия должна постепенно смещаться по мере изменения ориентации, в любой небольшой зоне изображения должен возникнуть радужный узор. Именно это и обнаружил Блэсдел в действительности. Примеров успешного применения этого метода пока еще мало, и нет возможности интерпретировать получаемые изображения в таких понятиях, как разрывы и реверсии, однако метод представляется многообещающим.
 Рис. 87. В этом эксперименте Г. Блэсдел использовал для обработки стриарной коры обезьяны окрашивание, чувствительное к электрическому напряжению. Обезьяне предъявлялись полосы последовательно изменяющейся ориентации. Изображение коры было получено с помощью телевизионной техники. С помощью компьютерной обработки изображений было получено окончательное изображение, на котором участкам, реагирующим на ту или иную ориентацию стимула, был приписан определенный цвет. В любой небольшой области коры ориентационные зоны проявляются в виде параллельных полос, так что в целом полный набор элементов, настроенных на разные ориентации, дает картину, окрашенную во все цвета радуги.
Рис. 87. В этом эксперименте Г. Блэсдел использовал для обработки стриарной коры обезьяны окрашивание, чувствительное к электрическому напряжению. Обезьяне предъявлялись полосы последовательно изменяющейся ориентации. Изображение коры было получено с помощью телевизионной техники. С помощью компьютерной обработки изображений было получено окончательное изображение, на котором участкам, реагирующим на ту или иную ориентацию стимула, был приписан определенный цвет. В любой небольшой области коры ориентационные зоны проявляются в виде параллельных полос, так что в целом полный набор элементов, настроенных на разные ориентации, дает картину, окрашенную во все цвета радуги.
Карты коры
Теперь, когда мы узнали кое-что о топографическом представлении в коре параметров ориентации и глазодоминантности, мы можем рассмотреть взаимосвязь между этими отображениями и проекциями зрительных полей. Обычно говорят, что сетчатка отображается на кору взаимно однозначно. Однако, учитывая то, что нам известно о рецептивных полях корковых клеток, можно с уверенностью сказать, что это утверждение, строго говоря, неверно, поскольку каждая клетка получает входные сигналы от тысяч палочек и колбочек, а ее рецептивное поле — далеко не точка. Отображение сетчатки на кору гораздо более сложно, чем любое обычное взаимно однозначное отображение. На рис. 88 я попытался показать распределение в коре тех областей, которые активируются под действием простого стимула (не следует смешивать это с рецептивным полем одиночного нейрона). Стимул здесь — это короткий отрезок линии с наклоном в 60° относительно вертикали, предъявленный только левому глазу. Мы предполагаем, что этот элемент поля зрения проецируется на область коры, выделенную прямоугольником с закругленными углами. В пределах этой области будут возбуждены только те полосы, которые связаны с левым глазом, а в этих полосах — только те участки, которые соответствуют ориентации в 60°. Все они на рисунке зачернены. Таким образом, линия, появившаяся в поле зрения, вызывает причудливую картину распределения возбужденных клеток коры в виде ряда узких полосок.
 Рис. 88. Наклонный отрезок прямой, предъявленный левому глазу (справа), мог бы вызывать распределение возбуждения в небольшой зоне стриарной коры, показанное слева. Возбужденные участки будут ограничены небольшой зоной коры — длинной и узкой в соответствии с формой стимула. В пределах само́й этой зоны возбуждение ограничено колонками глазодоминантности для левого глаза и только теми ориентационными колонками, которые соответствуют ориентации часовой стрелки в 2.00 или 8.00. Как видно, корковое отображение данного стимула не так просто!
Рис. 88. Наклонный отрезок прямой, предъявленный левому глазу (справа), мог бы вызывать распределение возбуждения в небольшой зоне стриарной коры, показанное слева. Возбужденные участки будут ограничены небольшой зоной коры — длинной и узкой в соответствии с формой стимула. В пределах само́й этой зоны возбуждение ограничено колонками глазодоминантности для левого глаза и только теми ориентационными колонками, которые соответствуют ориентации часовой стрелки в 2.00 или 8.00. Как видно, корковое отображение данного стимула не так просто!
Теперь становится понятно, насколько наивно представление о зеленом гомункулюсе, который сидит у нас в голове и рассматривает передаваемую в кору картину. На самом деле картина, которая создается в коре, имеет примерно такое же отношение к изображению внешнего мира, как электрические процессы в телекамере — к передаваемой внешней сцене. Если бы распределение корковой активности точно воспроизводило распределение яркости в поле зрения, то это означало бы, что на пути от сетчатки в кору не произошло ничего существенного. В этом случае действительно потребовался бы маленький зеленый человечек. Вряд ли можно представить себе, что в процессе эволюции было затрачено столько усилий на столь изящную группировку клеток в двух взаимонезависимых системах колонок, если бы это не обеспечивало какой-то пользы для животного. Пока мы не докопаемся до точной схемы связей, определяющей отображение сетчатки в коре, мы вряд ли сможем до конца понять принципы группировки клеток в коре. Сейчас мы можем только строить логичные догадки. Если мы предположим, что схема, приведенная в главе 4, соответствует реальности, то придется признать: то, что нужно для перехода от простых клеток к сложным или для создания чувствительности к концам линий или к направлению движения, — это определенного рода конвергенция множества клеток на одну клетку, причем у рецептивных полей промежуточных клеток должна быть одна и та же ориентация и примерно одно и то же положение. Пока у нас не было особых оснований предполагать, что клетка с определенной ориентацией рецептивного поля получает входы от клеток с рецептивными полями иной ориентации. (Я здесь несколько преувеличиваю: высказывались предположения, что клетки с разной оптимальной ориентацией стимулов могут быть соединены тормозными связями; в пользу этого имеются лишь косвенные данные, на мой взгляд, не очень убедительные, хотя их нельзя игнорировать.) Если это так, то почему бы не группировать вместе клетки, которые должны быть взаимно связаны? Альтернативный вариант едва ли целесообразен — легко представить себе, как трудно было бы обеспечить надлежащие связи между клетками, если эти клетки разбросаны в коре без учета их сходных свойств! Особенно много связей должно быть между клетками с одинаковой оптимальной ориентацией, и если бы такие клетки распределялись случайным образом, то сеть аксонов, необходимых для их соединения, заполнила бы все пространство между нейронами. Как бы то ни было, фактически они группируются вместе. Изложенные соображения относятся и к клеткам, сходным по глазодоминантности. Но если клетки со сходными свойствами упаковываются вместе, то почему, продвигая электрод, мы наблюдаем последовательное изменение ориентации небольшими шагами? И чем объясняется цикличность — почему, пройдя все возможные варианты, мы опять возвращаемся к исходной ориентации, а потом весь цикл повторяется снова? Казалось бы, можно было собрать вместе все клетки с ориентацией 30°, все клетки с ориентацией 42° и т.д.; и наконец, объединить все клетки с доминированием левого глаза и все — с доминированием правого глаза. Если бы мы знали, как устроена кора, то смогли бы предложить много ответов на эти вопросы. Сейчас выскажем одно предположение: возможно, что клетки с разной ориентацией действительно оказывают тормозящее воздействие друг на друга. Если нежелательно, чтобы какая-то клетка отвечала на другие ориентации, отличные от ее собственной, то нетрудно представить себе наличие тормозных связей, которые обостряют настройку данной клетки на определенную ориентацию. Тогда наблюдаемая картина — как раз то, что нам нужно: клетки с одинаковой ориентацией расположены ближе всего друг к другу, но не слишком далеко от клеток с иной, но очень сходной ориентацией; тогда тормозные связи будут достаточно короткими. Возможно другое предположение: если рассмотреть, какие входы от клеток слоя 4 (с круглыми полями) нужны для организации простой клетки с определенной оптимальной ориентацией, легко увидеть, что потребуется лишь небольшое изменение набора входов для того, чтобы получить близко расположенную простую клетку с близкой, но несколько иной ориентацией. Для этого достаточно добавить несколько новых входов, а несколько прежних убрать (рис. 89). Такого рода соображения позволили бы объяснить физическую близость расположения клеток со сходной ориентацией. Предметом следующей главы будет взаимосвязь между ориентацией, глазодоминантностью и проекцией поля зрения на кору. Рассмотрение этого вопроса может помочь нам понять, почему нужно так много колонок. Когда к этому добавится еще вопрос о топографии, соответствующая корковая организация будет представляться еще более сложной и интересной.
 Рис. 89. Показана группа клеток слоя 4 с концентрическими рецептивными полями, необходимая для создания простой клетки, отвечающей на наклонную линию с ориентацией «4.00 — 10.00». Вероятно, эта группа имеет общие клетки с группой, необходимой для реакции на линию с ориентацией «4.30 — 10.30», — для превращения одной группы в другую достаточно лишь несколько связей убрать, а несколько добавить.
Рис. 89. Показана группа клеток слоя 4 с концентрическими рецептивными полями, необходимая для создания простой клетки, отвечающей на наклонную линию с ориентацией «4.00 — 10.00». Вероятно, эта группа имеет общие клетки с группой, необходимой для реакции на линию с ориентацией «4.30 — 10.30», — для превращения одной группы в другую достаточно лишь несколько связей убрать, а несколько добавить.
6. «Увеличение» и модули
В предыдущей главе я подчеркивал, что первичная зрительная кора кажется морфологически однородной как невооруженному глазу, так и под микроскопом при использовании большинства обычных методов окрашивания. Однако при более тщательном изучении выяснилось, что однородна и топография колонок глазодоминантности: период чередования зон доминирования левого и правого глаза остается на удивление постоянным от проекции центральной ямки (точки фиксации) до дальней периферии бинокулярной части поля зрения. При помощи метода с инъекцией дезоксиглюкозы мы выявили также однородность топографии ориентационных колонок. Факт анатомической однородности коры мы вначале восприняли как неожиданность, так как с функциональной точки зрения эта область явно неоднородна по двум важным параметрам. Во-первых, как уже говорилось в главе 3, рецептивные поля ганглиозных клеток сетчатки в зоне центральной ямки и около нее гораздо меньше, чем на периферии. Размеры рецептивного поля типичной сложной клетки в верхнем слое коры в области проекции центральной ямки составляют примерно 0,25×0,5°. Если же рассматривать те части коры, где представлена периферия сетчатки (80–90° от центральной ямки), то здесь чаще всего встречаются рецептивные поля размером 2×4°. Если сравнивать соответствующие площади, то отношение между ними можно оценить как 1:10 или даже 1:30. Второй вид неоднородности связан с так называемым «увеличением». Это понятие, введенное в 1961 году П. Дэниелом и Д. Уиттериджем, означает расстояние между двумя точками коры, в которые проецируются две точки поля зрения, разделенные расстоянием в 1°. Если идти от центральной ямки к периферии, корковая проекция одного и того же углового расстояния будет становиться все меньше и меньше, т.е. «увеличение» будет уменьшаться. Если сместить стимул на 1° около точки фиксации, то его проекция в коре сдвинется примерно на 6 мм. Если же поместить этот стимул в зону, отстоящую от точки фиксации на 90°, то сдвиг в поле зрения на 1° будет соответствовать сдвигу вдоль поверхности коры всего лишь на 0,15 мм. Таким образом, «увеличение» в центре сетчатки примерно в 36 раз больше, чем на периферии. Обе рассмотренные неоднородности связаны с одним и тем же обстоятельством — с уменьшением остроты зрения по мере удаления от центра сетчатки к ее периферии. Попробуйте, например, фиксировать взором какую-нибудь букву в самом начале строки и попытайтесь определить, какая буква или слово находится в самом ее конце. Или фиксируйте взглядом букву п в начале слова прогрессивно. Вряд ли вам удастся опознать букву о в конце слова. Вероятно, вам трудно будет опознать также в и н перед последней буквой. Для того чтобы обеспечить высокое разрешение в центральной ямке, зрительной системе требуется много корковых клеток на единицу площади в проекции этого участка, причем нужно, чтобы каждая такая клетка имела очень небольшое рецептивное поле. Рис. 90. В этой главе речь пойдет о «модулях». Один модуль по размерам примерно соответствует изображенному здесь срезу зрительной коры, окрашенному по Гольджи. Этот метод окраски выявляет лишь очень малую долю всех нервных клеток, но если уж клетка оказывается окрашенной, то она видна вся полностью или почти вся: на таком срезе можно рассмотреть тело клетки, ее дендриты и аксон.
Рис. 90. В этой главе речь пойдет о «модулях». Один модуль по размерам примерно соответствует изображенному здесь срезу зрительной коры, окрашенному по Гольджи. Этот метод окраски выявляет лишь очень малую долю всех нервных клеток, но если уж клетка оказывается окрашенной, то она видна вся полностью или почти вся: на таком срезе можно рассмотреть тело клетки, ее дендриты и аксон.
Разброс и смещение рецептивных полей
Учитывая сказанное выше, как можно объяснить анатомическую однородность коры? Для этого придется подробнее рассмотреть, что́ происходит с положением рецептивных полей нейронов, встречающихся на пути продвижения электрода. Если мы будем вводить электрод в стриарную кору точно под прямым углом к ее поверхности, то окажется, что все эти рецептивные поля расположены почти в одном и том же участке поля зрения, но не совпадают полностью — от клетки к клетке положение рецептивного поля несколько меняется. Однако эти изменения кажутся случайными и не очень велики, так что рецептивные поля каждых двух последовательных клеток значительно перекрываются (этот факт отражен на рис. 91). В каждом данном слое коры размеры рецептивных полей относительно постоянны, однако при переходе к другому слою они заметно меняются — от очень малых в слое 4C до больших в слоях 5 и 6. В пределах одного слоя коры площадь, занимаемая десятью или двенадцатью последовательно зарегистрированными полями, из-за случайного разброса будет примерно в 2–4 раза больше, чем площадь рецептивного поля одной клетки. Назовем ту область, которую занимает вся совокупность рецептивных полей отдельных клеток в одном слое и в одной точке коры составным рецептивным полем данной точки. В зависимости от слоя коры величина составного поля варьирует — например, в слое 3 она равна около 30 угловых минут для центральной ямки и примерно 7–8° для дальней периферии.
 Рис. 91. Картирование этих девяти рецептивных полей было проведено при однократном прохождении микроэлектрода через стриарную кору кошки перпендикулярно к ее поверхности. При таком продвижении электрода наблюдаются случайные отклонения в положении рецептивных полей, а также в их величине, но не отмечается какой-либо тенденции к их систематическому смещению.
Рис. 91. Картирование этих девяти рецептивных полей было проведено при однократном прохождении микроэлектрода через стриарную кору кошки перпендикулярно к ее поверхности. При таком продвижении электрода наблюдаются случайные отклонения в положении рецептивных полей, а также в их величине, но не отмечается какой-либо тенденции к их систематическому смещению.
Теперь предположим, что мы вводим и продвигаем электрод горизонтально в пределах одного слоя коры, скажем слоя 3. И здесь при последовательной регистрации ответов клеток мы обнаружим, что положение рецептивных полей варьирует случайным образом, однако в этих беспорядочных сдвигах мы заметим тенденцию к смещению в определенном направлении. Разумеется, это направление можно предсказать исходя из топографической проекции поля зрения в кору. Сейчас нас будет интересовать величина смещения, которая наблюдается при сдвиге электрода на 1 миллиметр по горизонтали. Если учесть то, что было сказано выше относительно различий в «увеличении», ясно, что расстояние, измеряемое в поле зрения, будет зависеть от того, где именно в коре мы продвигаем электрод — в области проекции центральной ямки, в периферийной области или где-то в промежуточной зоне. Скорость смещения рецептивных полей в поле зрения будет далеко не одинаковой. Оказывается, однако, что эта скорость будет очень близка к постоянной, если сделать поправку на размер самих рецептивных полей. Смещение в любом месте коры на 1 миллиметр соответствует такому сдвигу в поле зрения, который равен примерно половине поперечника составного рецептивного поля (т.е. суммарного поля, образуемого наложением рецептивных полей всех клеток, находящихся под данной точкой поверхности коры. Таким образом, чтобы полностью выйти из одного участка поля зрения и перейти в другой, следующий, потребуется смещение примерно в 2 миллиметра (это показано на рис. 92). То же оказывается справедливым для всех точек поля 17, где мы отводили электрическую активность. В области центральной ямки рецептивные поля очень малы, однако невелик и сдвиг в поле зрения, который соответствует смещению вдоль коры на 2 миллиметра. Напротив, на периферии как размеры рецептивных полей, так и соответствующие смещения в поле зрения гораздо больше (рис. 93).
 Рис. 92. При продвижении электрода в коре кошки на большое расстояние параллельно поверхности положение рецептивных полей встречающихся клеток изменяется. В данном случае электрод прошел 3 мм и встретился примерно с 60 клетками (их слишком много, чтобы показать на этом рисунке). Здесь отмечены положения только четырех или пяти рецептивных полей, карты для которых составлялись в начале прохождения каждого миллиметра (примерно десятой его части), а остальные поля оставлялись без внимания. В верхней части рисунка показаны рецептивные поля клеток, встретившихся на четырех этапах проходки (0, 1, 2 и 3). Каждая такая группа полей заметно смещалась в поле зрения вправо относительно предыдущей группы. Рецептивные поля клеток группы 2 не перекрываются с полями группы 0, а поля группы 3 — с полями группы 1. В обоих случаях расстояние вдоль коры составляло 2 миллиметра.
Рис. 92. При продвижении электрода в коре кошки на большое расстояние параллельно поверхности положение рецептивных полей встречающихся клеток изменяется. В данном случае электрод прошел 3 мм и встретился примерно с 60 клетками (их слишком много, чтобы показать на этом рисунке). Здесь отмечены положения только четырех или пяти рецептивных полей, карты для которых составлялись в начале прохождения каждого миллиметра (примерно десятой его части), а остальные поля оставлялись без внимания. В верхней части рисунка показаны рецептивные поля клеток, встретившихся на четырех этапах проходки (0, 1, 2 и 3). Каждая такая группа полей заметно смещалась в поле зрения вправо относительно предыдущей группы. Рецептивные поля клеток группы 2 не перекрываются с полями группы 0, а поля группы 3 — с полями группы 1. В обоих случаях расстояние вдоль коры составляло 2 миллиметра.
 Рис. 93. У макака рецептивные поля клеток верхних слоев коры увеличиваются по мере удаления от центральной ямки (которой соответствует отметка 0°). В таком же отношении увеличивается и расстояние между центрами рецептивных полей тех клеток, расстояние между которыми в коре составляет 2 миллиметра.
Рис. 93. У макака рецептивные поля клеток верхних слоев коры увеличиваются по мере удаления от центральной ямки (которой соответствует отметка 0°). В таком же отношении увеличивается и расстояние между центрами рецептивных полей тех клеток, расстояние между которыми в коре составляет 2 миллиметра.
Функциональные единицы коры
Нам приходится заключить, что любой участок первичной зрительной коры величиной примерно 2×2 миллиметра должен иметь все механизмы, необходимые для полной обработки определенного участка поля зрения, который очень мал в области центральной ямки и гораздо больше на периферии. Такой участок, получающий, вероятно, десятки тысяч входных волокон от наружного коленчатого тела, перерабатывает поступающую по ним информацию и посылает выходные сигналы по волокнам, чувствительным к ориентации стимула, его движению и т.д., объединяя информацию от обоих глаз. Все участки производят сходные операции над сигналами, поступающими примерно по одному и тому же числу волокон. Информация, получаемая отдельным участком, весьма детальна, если поступает с малой площади в области центральной ямки, и менее подробна, когда приходит с большей площади с периферии сетчатки. Выходные сигналы такого участка вырабатываются без всякого учета того, какой по величине участок поля зрения здесь анализируется и с какой степенью детализации. Набор механизмов для переработки информации в таких участках в основном один и тот же. Это позволяет объяснить ту однородность, которую мы видим и невооруженным глазом, и при микроскопическом исследовании. Тот факт, что сдвига вдоль коры на 2 мм как раз достаточно, чтобы попасть на проекцию нового участка сетчатки, означает, что какие бы локальные операции ни выполнялись в коре, все они должны выполняться в пределах блока размером 2×2 мм. Очевидно, меньший участок не сможет всесторонне анализировать соответственно меньшую площадку поля зрения, так как для такого анализа нужны все элементы 2-миллиметрового блока. Это ясно уже из рассмотрения данных о положении и размерах рецептивных полей. Однако нам надо будет подробнее обсудить, что означают слова «анализ» и «операция». Можно начать с рассмотрения параметра ориентации линии. В любом участке поля зрения, даже самом малом, должны быть учтены все возможные ориентации. Если при анализе какого-то участка соответствующим корковым блоком величиной 2 мм в этом блоке не окажется элемента, реагирующего на ориентацию +45°, то наличие такого элемента в других блоках коры не поможет, так как другие блоки имеют дело с другими участками поля зрения. Однако, к счастью, ширина ориентационных полос в коре достаточно мала — 0,05 мм, и набор таких полос для всех ориентаций от 0 до 180° с шагом в 10° дважды укладывается на участке коры шириной 2 мм. Так же обстоит дело и с глазодоминантностью — ширина соответствующих колонок 0,5 мм, поэтому 2-миллиметрового блока более чем достаточно для полного анализа. Таким образом, блоки поперечником в 2 мм обладают, по-видимому, полным набором необходимых механизмов.
 Рис. 94. Мы называем эту схему «нашей моделью из ледяных кубиков». Схема показывает, как разбита кора одновременно на два вида пластин — для глазодоминантности (для правого и левого глаза) и для ориентации. Эту модель не следует воспринимать буквально — ни одна из этих систем пластин не бывает такой регулярной, особенно ориентационные пластины, которые далеки от того, чтобы располагаться параллельно и иметь прямые границы. Более того, они, по-видимому, не пересекаются под каким-то определенным углом и уж во всяком случае не образуют ортогональную систему вроде показанной на рисунке.
Рис. 94. Мы называем эту схему «нашей моделью из ледяных кубиков». Схема показывает, как разбита кора одновременно на два вида пластин — для глазодоминантности (для правого и левого глаза) и для ориентации. Эту модель не следует воспринимать буквально — ни одна из этих систем пластин не бывает такой регулярной, особенно ориентационные пластины, которые далеки от того, чтобы располагаться параллельно и иметь прямые границы. Более того, они, по-видимому, не пересекаются под каким-то определенным углом и уж во всяком случае не образуют ортогональную систему вроде показанной на рисунке.
Следует добавить, что блоки величиной 2 мм — это особенность не столько всего поля 17 коры, сколько слоя 3 поля 17. Если обратиться к слоям 5 и 6, то там размеры рецептивных полей клеток и их разброс вдвое больше; поэтому для всестороннего анализа в слоях 5 и 6 и образования рецептивных полей большей величины и с более сложными свойствами, по-видимому, потребуются блоки с поперечником в 4 мм. С другой стороны, в слое 4C рецептивные поля и их разброс гораздо меньше и соответственно размеры блоков ближе к 0,1–0,2 мм. Однако общая идея отдельных функциональных блоков остается прежней независимо от того факта, что любая заданная область поля зрения обслуживается набором локальных механизмов, размещенных в нескольких слоях коры, т.е. что кора — это совокупность нескольких систем, объединенных в одно целое. Все эти соображения помогают нам понять, почему колонки не могут быть гораздо более крупными. В блок размером 2×2 мм нужно поместить все механизмы, выделяющие нужные переменные в стимулах. До сих пор мы говорили об ориентации и глазодоминантности, однако на самом деле на двумерную поверхность отображаются не две, а большее число входных переменных. Для этого в качестве основных параметров выбраны две переменные, определяющие координаты в поле зрения (расстояния от центральной ямки по горизонтали и по вертикали). На полученной координатной сетке в виде более тонкой мозаики дополнительно отображены и другие переменные, такие как ориентация стимула и глазодоминантность. Мы будем называть участок коры с поперечником 2 мм модулем. Лично мне этот термин кажется не вполне подходящим, в частности, потому, что он вызывает слишком конкретное представление о небольшом прямоугольном блоке электронного устройства, который вместе с сотнями других таких же блоков смонтирован на общем шасси. Однако в определенной мере мы имели в виду именно эту ассоциацию, но более обобщенного характера. Во-первых, выделенные нами функциональные модули ориентационной системы таковы, что границы между смежными модулями условны. Например, можно считать, что модуль начинается с клеток, выделяющих вертикальную ориентацию, и заканчивается следующей группой клеток, которые тоже выделяют вертикальную ориентацию; за начало можно принять и любую другую ориентацию, лишь бы в каждом модуле все ориентации были представлены хотя бы по одному разу. Так же обстоит дело и с глазодоминантностью — модуль может начинаться с того места, где доминирует правый глаз, с того, где доминирует левый, или с середины колонки, лишь бы в конечном счете в модуль вошли два участка глазодоминантности, по одному для каждого глаза. Во-вторых, как мы уже упоминали, величина модуля будет зависеть от того, какой из слоев коры мы рассматриваем. Тем не менее термин «модуль» должен ассоциироваться с системой примерно из 500–1000 миниатюрных устройств, которые все взаимозаменяемы (при условии, что можно было бы воспроизвести около 10 000 входных и, вероятно, около 50 000 выходных связей). Необходимо тут же добавить, что никто, конечно, не думает, что кора совершенно однородна, — от проекции центральной ямки до участков, отображающих дальнюю периферию. Помимо остроты зрения по мере удаления от центра сетчатки изменяются и другие параметры зрительного восприятия; ухудшается, например, цветовое зрение, хотя, возможно, и не очень резко, если компенсировать уменьшение коэффициента «усиления» (т.е. увеличивать размеры объектов по мере удаления их от точки фиксации). Движение, равно как и очень слабые источники света, по всей вероятности, легче замечается на периферии поля зрения. Бинокулярное зрение должно ухудшаться, так как в зонах радиусом от 20 до 80° колонки глазодоминантности для ипсилатерального глаза становятся все уже и уже, а для контралатерального глаза — все шире и шире. На расстояниях больше 80° от центральной ямки ипсилатеральные колонки полностью исчезают и корковые механизмы становятся «монокулярными». Ввиду этих, а также и других различий, которые, несомненно, имеются, соответствующие корковые структуры должны быть в чем-то неоднородными. Поэтому корковые модули, вероятно, не все совершенно одинаковы.
Деформация коры
Если сравнивать кору с сетчаткой, то можно получить более полное представление о конфигурации коры. Глаз представляет собой сферу, поскольку это чисто оптический инструмент. Поэтому форма сетчатки тоже сферическая. В фотоаппарате пленка может быть плоской, так как апертура линзы составляет в среднем около 30°. В широкоугольной камере типа «рыбий глаз» апертура гораздо больше, однако на периферии изображение искажается. Конечно, фотографии сферической формы были бы неудобны — плоские фотоснимки гораздо удобнее хранить. Что касается глаза, то его шаровидная форма представляется идеальной, так как шар — тело компактное, и в то же время он может легко вращаться в глазнице (если бы глаз имел форму куба, то возникли бы затруднения). А поскольку глаз имеет сферическую форму, то угловой масштаб изображения зрительного поля на сетчатке остается везде постоянным — число градусов, приходящееся на 1 мм сетчатки, на всей сетчатке одинаково. У человека оно составляет 3,5°. Как уже говорилось, центры рецептивных полей ганглиозных клеток в центральной ямке и поблизости от нее очень малы, но они увеличиваются по мере удаления от нее. Поэтому мы не должны удивляться тому, что в центральной области сетчатки в каждом миллиметре должно быть гораздо больше ганглиозных клеток, чем в зоне дальней периферии. Действительно, вблизи центральной ямки эти клетки уложены несколькими слоями, тогда как на периферии сетчатки располагаются настолько редко, что визуально не образуют даже одного непрерывного слоя (см. микрофотографии на рис. 95). Поскольку сетчатка должна иметь сферическую форму, то ее слои не могут быть однородными. Возможно, это одна из причин того, что в сетчатке происходит лишь весьма ограниченная обработка информации; иначе потребовалось бы очень большое утолщение ее слоев в центральном участке. В этом смысле кора обладает бо́льшими возможностями. В отличие от сетчатки она может не быть сферической, а может просто растягиваться в области проекции центральной ямки по сравнению с проекцией периферии. Это растяжение, по-видимому, осуществляется в такой степени, чтобы толщина коркового слоя (так же как ширина колонок и другие параметры) была повсюду одинаковой. Какое влияние это оказывает на общую форму коры? Хотя я неоднократно характеризовал кору как «слой», это не означает, что она должна быть плоской. Если бы не было никаких искажений конфигурации, стриарная кора имела бы сферическую форму — такую же как у глазного яблока. В этом смысле она похожа на поверхность Земли — без искажений, связанных с рельефом, поверхность Земли имела бы строго сферическую форму. Правда, на стриарную кору одного полушария отображается лишь около половины задней поверхности глаза, т.е. примерно четверть всей сферы.
 Рис. 95. В отличие от слоев коры слои сетчатки далеко не одинаковы по толщине в различных ее участках. Как у обезьяны, так и у человека слой ганглиозных клеток вблизи центральной ямки (нижний слой на верхней микрофотографии) имеет толщину в 8–10 клеток, в то время как на дальней периферии сетчатки, на расстоянии 70–80° от ее центра (нижняя микрофотография) ганглиозные клетки не образуют даже одного сплошного слоя. Это не должно нас удивлять, поскольку центры рецептивных полей ганглиозных клеток около центральной ямки очень малы, а на периферии они увеличиваются. В центре сетчатки на единицу площади приходится гораздо больше ганглиозных клеток.
Рис. 95. В отличие от слоев коры слои сетчатки далеко не одинаковы по толщине в различных ее участках. Как у обезьяны, так и у человека слой ганглиозных клеток вблизи центральной ямки (нижний слой на верхней микрофотографии) имеет толщину в 8–10 клеток, в то время как на дальней периферии сетчатки, на расстоянии 70–80° от ее центра (нижняя микрофотография) ганглиозные клетки не образуют даже одного сплошного слоя. Это не должно нас удивлять, поскольку центры рецептивных полей ганглиозных клеток около центральной ямки очень малы, а на периферии они увеличиваются. В центре сетчатки на единицу площади приходится гораздо больше ганглиозных клеток.
 Рис. 96. Расправленная стриарная кора по форме напоминала бы грушу. Она имела бы вид четверти сферы, если бы все части сетчатки были представлены в ней пропорционально. На самом деле, однако, форма коры значительно искажена из-за несоразмерно увеличенного отображения центральной области сетчатки. Дальняя периферия (примерно в 90° от центра) отображена так слабо, что полукруг, соответствующий 90° (крайний справа), оказывается очень небольшим.
Рис. 96. Расправленная стриарная кора по форме напоминала бы грушу. Она имела бы вид четверти сферы, если бы все части сетчатки были представлены в ней пропорционально. На самом деле, однако, форма коры значительно искажена из-за несоразмерно увеличенного отображения центральной области сетчатки. Дальняя периферия (примерно в 90° от центра) отображена так слабо, что полукруг, соответствующий 90° (крайний справа), оказывается очень небольшим.
Растягиваясь таким образом, чтобы сохранять постоянную толщину и все же иметь возможность переработать особенно обильную информацию от множества ганглиозных клеток центральной ямки, кора приобретает форму, отличную от сферической. Если развернуть и разгладить все складки коры, форма ее не окажется ни сферической, ни плоской. В действительности кора будет тогда иметь вид сильно искаженного сегмента сферы, составляющего четвертую ее часть; по форме она скорее будет напоминать грушу или яйцо. Такой результат предсказывали в 1962 году Дэниел и Уиттеридж, экспериментально определявшие зависимость «увеличения» в поле 17 от расстояния до проекции центральной ямки (об этом говорилось на с. 0127). Из полученных данных они вычислили трехмерную форму поверхности коры, а затем по серии последовательных гистологических срезов сделали резиновую модель. После этого они на самом деле расправили эту модель, тем самым подтвердив свое предсказание относительно грушевидной формы коры. Полученная форма показана на рис. 96. До этого никто не задавался вопросом, какую форму будет иметь зрительная кора, если расправить ее складки. Насколько мне известно, никто также не сознавал, что любая область коры должна иметь какую-то определенную форму, логически вытекающую из функций этой области. По-видимому, складки коры, которые для выяснения истинной формы нужно разгладить (без растяжения и разрывов), образуются из-за того, что большой сегмент коры должен был быть «скомкан», чтобы поместиться в ограниченном пространстве черепной коробки. При этом характер складок может быть не совсем случайным; какие-то детали, вероятно, определяются тем, что межкортикальные связи должны быть по возможности короче. Если обратиться к соматосенсорной коре, то здесь сложность топографического отображения может доходить до крайней степени. Например, область коры, соответствующая коже руки, по конфигурации должна быть похожа на перчатку, причем с определенными искажениями, чтобы обеспечить бо́льшую чувствительность кончиков пальцев по сравнению с ладонью и тыльной стороной кисти. Такое искажение аналогично увеличению корковой проекции центральной ямки по сравнению с периферией сетчатки в соответствии с различием в остроте зрения. Будет ли корковая проекция руки действительно напоминать перчатку, если мы сделаем резиновую модель и будем ее постепенно раздувать, чтобы расправить искусственные складки? Вероятно, нет. Картирование соматосенсорной коры оказалось чрезвычайно сложной задачей. Результаты, полученные до сих пор, позволяют думать, что форма непрерывной проекции была бы слишком причудливой; на самом деле соматосенсорная кора как будто ножницами разрезана на множество небольших, более «удобных» участков, которые соединены наподобие кусков лоскутного одеяла так, что получается некоторое приближение к плоской поверхности.
7. Мозолистое тело и стереоскопическое зрение
Мозолистое тело представляет собой мощный пучок миелинизированных волокон, соединяющих два полушария мозга. Стереоскопическое зрение (стереопсис) — это способность воспринимать глубину пространства и оценивать удаленность предметов от глаз. Эти две вещи не особенно тесно связаны друг с другом, однако известно, что небольшая часть волокон мозолистого тела все же играет некоторую роль в стереопсисе. Оказалось удобным включить обе эти темы в одну главу, так как при их рассмотрении придется учитывать одну и ту же особенность устройства зрительной системы, а именно то, что в хиазме имеются как перекрещенные, так и неперекрещенные волокна зрительного нерва (см. рис. 35).Мозолистое тело
Мозолистое тело (по-латыни corpus callosum) — это самый крупный пучок нервных волокон во всей нервной системе. По приближенной оценке в нем насчитывается около 200 млн. аксонов. Истинное число волокон, вероятно, еще больше, так как приведенная оценка основана на данных обычной световой, а не электронной микроскопии. Это число несравнимо с числом волокон в каждом зрительном нерве (1,5 млн.) и в слуховом нерве (32 000). Площадь поперечного сечения мозолистого тела составляет около 700 мм2, тогда как у зрительного нерва она не превышает нескольких квадратных миллиметров. Мозолистое тело вместе с тонким пучком волокон, называемым передней комиссурой, соединяет два полушария мозга (рис. 98 и 99). Термин комиссура означает совокупность волокон, соединяющих две гомологичные нервные структуры, расположенные в левой и правой половинах головного или спинного мозга. Мозолистое тело тоже иногда называют большой комиссурой мозга.
 Рис. 97. На этой стереопаре изображена галерея в Новом колледже в Оксфорде. После того как была сделана сдвинули на 7,5 см влево и сделали левую фотографию.
Рис. 97. На этой стереопаре изображена галерея в Новом колледже в Оксфорде. После того как была сделана сдвинули на 7,5 см влево и сделали левую фотографию.
 Рис. 98. Головной мозг человека в разрезе, проходящем по срединной плоскости. Виден толстый, дугообразный в поперечном сечении пучок аксонов — мозолистое тело.
Рис. 98. Головной мозг человека в разрезе, проходящем по срединной плоскости. Виден толстый, дугообразный в поперечном сечении пучок аксонов — мозолистое тело.
 Рис. 99. Здесь мозг показан сверху. Часть правого полушария срезана, и виден пучок волокон мозолистого тела, соединяющий все участки двух полушарий.
Рис. 99. Здесь мозг показан сверху. Часть правого полушария срезана, и виден пучок волокон мозолистого тела, соединяющий все участки двух полушарий.
Примерно до 1950 года роль мозолистого тела была совершенно неизвестна. В редких случаях наблюдается врожденное отсутствие (аплазия) мозолистого тела. Это образование может также быть частично или полностью перерезано во время нейрохирургической операции, что делается намеренно — в одних случаях при лечении эпилепсии (чтобы судорожный разряд, возникающий в одном полушарии мозга, не мог распространиться на другое полушарие), в других случаях для того, чтобы добраться сверху до глубоко расположенной опухоли (если, например, опухоль находится в гипофизе). По наблюдениям невропатологов и психиатров, после такого рода операций не возникает никаких расстройств психики. Кто-то даже высказал мысль (хотя вряд ли всерьез), что единственная функция мозолистого тела состоит в том, чтобы удерживать два полушария мозга вместе. Вплоть до 1950-х годов мало что было известно о деталях распределения связей в мозолистом теле. Очевидно было, что мозолистое тело соединяет два полушария, и на основании данных, полученных довольно грубыми нейрофизиологическими методами, считали, что в стриарной коре волокна мозолистого тела связывают в точности симметричные участки двух полушарий. В 1955 году Рональд Майерс, аспирант психолога Роджера Сперрииз Чикагского университета, впервые провел эксперимент, в котором удалось выявить некоторые функции этого огромного волокнистого тракта. Майерс занимался обучением кошек, помещенных в ящик с двумя поставленными рядом экранами, на которые можно было проецировать различные изображения, например круг на один экран и квадрат — на другой. Кошку обучали упираться носом в тот экран, на котором было изображение круга, и игнорировать другой — с изображением квадрата. Правильные ответы подкреплялись пищей, а за ошибочные ответы кошек слегка наказывали — включался громкий звонок, и кошку не грубо, но решительно оттаскивали от экрана. Таким методом за несколько тысяч повторений кошку удается довести до уровня надежного различения фигур. (Кошки обучаются медленно; например, голубям для обучения в аналогичной задаче требуется от нескольких десятков до нескольких сотен повторений, а человека вообще можно научить сразу, дав ему словесную инструкцию. Такая разница кажется несколько странной — ведь у кошки головной мозг во много раз больше, чем у голубя.) Нет ничего удивительного в том, что кошки Майерса научались ничем не хуже решать эту задачу и в том случае, когда один глаз животного был закрыт маской. Неудивительно и то, что если обучение такой задаче, как выбор треугольника или квадрата, проводилось лишь с одним открытым глазом — левым, а при проверке левый глаз закрывали и открывали правый, то точность различения оставалась прежней. Нас это не удивляет потому, что мы сами легко можем решить аналогичную задачу. Легкость решения подобных задач понятна, если учесть анатомию зрительной системы. Каждое полушарие получает входные сигналы от обоих глаз. Как мы уже говорили в главе 4, большая часть клеток в поле 17 тоже имеет входы от обоих глаз. Майерс создал более интересную ситуацию, произведя продольную перерезку хиазмы по средней линии. Таким образом, он перерезал перекрещивающиеся волокна и сохранил в целости неперекрещивающиеся (эта операция требует от хирурга определенного навыка). В результате такой перерезки левый глаз животного оказался соединен только с левым полушарием, а правый — только с правым. Идея эксперимента заключалась в том, чтобы обучать кошку, используя левый глаз, а на «экзамене» адресовать стимул правому глазу. Если кошка сможет правильно решать задачу, то это будет означать, что необходимая информация передается из левого полушария в правое по единственному известному пути — через мозолистое тело. Итак, Майерс произвел продольную перерезку хиазмы, обучил кошку с одним открытым глазом, а затем устроил проверку, открыв другой глаз и закрыв первый. В этих условиях кошки по-прежнему успешно решали задачу. Наконец, Майерс повторил эксперимент на животных, у которых предварительно были перерезаны и хиазма, и мозолистое тело. На этот раз кошки задачу не решили. Таким образом, Майерс опытным путем установил, что мозолистое тело действительно выполняет какие-то функции (хотя вряд ли можно было думать, что оно существует только для того, чтобы отдельные люди или животные с перерезанной зрительной хиазмой могли решать определенные задачи с использованием одного глаза после обучения с использованием другого).
Изучение физиологии мозолистого тела
Одно из первых нейрофизиологических исследований в этой области было проведено спустя несколько лет после экспериментов Майерса Д. Уиттериджем, работавшим тогда в Эдинбурге. Уиттеридж рассудил, что нет особого смысла в том, чтобы пучки нервных волокон соединяли гомологичные зеркально-симметричные участки полей 17. Действительно, не видно никаких причин для того, чтобы нервная клетка в левом полушарии, связанная с какими-то точками в правой половине поля зрения, соединялась с клеткой в правом полушарии, связанной с симметричным участком левой половины поля зрения. Для проверки своих предположений Уиттеридж перере́зал зрительный тракт на правой стороне мозга позади хиазмы и тем самым перекрыл входным сигналам путь в правую затылочную долю; но это, конечно, не исключало передачу туда сигналов из левой затылочной доли через мозолистое тело (рис. 100). Затем Уиттеридж стал включать световой стимул и регистрировать металлическим электродом электрическую активность с поверхности коры. Он действительно получил в своем опыте ответы, однако они возникали только на внутренней границе поля 17, т.е. в зоне, получающей входные сигналы от длинной, узкой вертикальной полоски в середине поля зрения: при стимуляции маленькими пятнышками света ответы появлялись только тогда, когда свет вспыхивал на вертикальной средней линии или поблизости от нее. Если кору противоположного полушария охлаждали, тем самым временно подавляя ее функцию, ответы прекращались; к этому же приводило и охлаждение мозолистого тела. Тогда стало ясно, что мозолистое тело не может связывать всё поле 17 левого полушария со всем полем 17 правого полушария, а связывает только небольшие участки этих полей, где находятся проекции вертикальной линии в середине поля зрения. Подобный результат можно было предвидеть исходя из ряда анатомических данных. Только один участок поля 17, расположенный очень близко к границе с полем 18, посылает аксоны через мозолистое тело в другое полушарие, и большая часть их, по-видимому, оканчивается в поле 18 около границы с полем 17. Если мы предположим, что входы в кору от НКТ точно соответствуют контралатеральным частям поля зрения (а именно левое полуполе отображается в коре правого полушария, а правое — в коре левого), то наличие связей между полушариями через мозолистое тело должно приводить в итоге к тому, что каждое полушарие будет получать сигналы от области несколько большей, чем половина поля зрения. Иными словами, за счет связей через мозолистое тело произойдет перекрывание полуполей, проецируемых в два полушария. Именно это мы и обнаружили. С помощью двух электродов, введенных в область коры у границы полей 17 и 18 в каждом из полушарий, нам нередко удавалось регистрировать активность клеток, рецептивные поля которых взаимно перекрывались на несколько угловых градусов.
 Рис. 100. Уиттеридж в своем эксперименте произвел перерезку правого зрительного тракта. Теперь для того, чтобы информация поступала в зрительную кору правого полушария, она должна сначала попасть в зрительную кору левого полушария, а затем пройти в другое полушарие через мозолистое тело. Охлаждение какого-либо участка этого пути блокирует передачу нервных импульсов.
Рис. 100. Уиттеридж в своем эксперименте произвел перерезку правого зрительного тракта. Теперь для того, чтобы информация поступала в зрительную кору правого полушария, она должна сначала попасть в зрительную кору левого полушария, а затем пройти в другое полушарие через мозолистое тело. Охлаждение какого-либо участка этого пути блокирует передачу нервных импульсов.
Т. Визел и я вскоре произвели микроэлектродные отведения непосредственно от той зоны мозолистого тела (в самой задней его части), где имеются волокна, связанные со зрительной системой. Мы нашли, что почти все волокна, которые мы могли активировать зрительными стимулами, отвечали в точности так же, как и обычные нейроны поля 17, т.е. проявляли свойства как простых, так и сложных клеток, избирательно чувствительных к ориентации стимула и обычно отвечавших на стимуляцию обоих глаз. Во всех этих случаях рецептивные поля располагались очень близко к средней вертикали ниже или выше (либо на уровне) точки фиксации, как показано на рис. 101.
 Рис. 101. Рецептивные поля волокон мозолистого тела лежат очень близко к средней вертикали поля зрения. Все эти поля были выявлены путем регистрации активности десяти волокон у одной кошки.
Рис. 101. Рецептивные поля волокон мозолистого тела лежат очень близко к средней вертикали поля зрения. Все эти поля были выявлены путем регистрации активности десяти волокон у одной кошки.
 Рис. 102. Берлукки и Риццолатти в своем эксперименте изящно продемонстрировали не только функцию зрительной части волокон мозолистого тела, но и высокую специфичность межполушарных связей между клетками, сходными по ориентации и расположению рецептивных полей. Берлукки и Риццолатти перерезали у кошки хиазму по средней линии, так что левый глаз стал передавать информацию только в левое полушарие, причем эта информация касалась только правой половины поля зрения. Точно так же правый глаз стал снабжать информацией только правое полушарие, и эта информация относилась только к левой половине поля зрения. После перерезки регистрировались ответы тех клеток, рецептивные поля которых в нормальных условиях пересекали бы среднюю вертикаль. Оказалось, что рецептивные поля таких клеток расщепляются по этой вертикали; правая часть связана теперь с левым глазом, а левая — с правым глазом.
Рис. 102. Берлукки и Риццолатти в своем эксперименте изящно продемонстрировали не только функцию зрительной части волокон мозолистого тела, но и высокую специфичность межполушарных связей между клетками, сходными по ориентации и расположению рецептивных полей. Берлукки и Риццолатти перерезали у кошки хиазму по средней линии, так что левый глаз стал передавать информацию только в левое полушарие, причем эта информация касалась только правой половины поля зрения. Точно так же правый глаз стал снабжать информацией только правое полушарие, и эта информация относилась только к левой половине поля зрения. После перерезки регистрировались ответы тех клеток, рецептивные поля которых в нормальных условиях пересекали бы среднюю вертикаль. Оказалось, что рецептивные поля таких клеток расщепляются по этой вертикали; правая часть связана теперь с левым глазом, а левая — с правым глазом.
Пожалуй, наиболее изящной нейрофизиологической демонстрацией роли мозолистого тела стала работа Дж. Берлукки и Дж. Риццолатти из г. Пизы, выполненная в 1968 году. Перерезав зрительную хиазму по средней линии, они регистрировали ответы в поле 17 вблизи границы с полем 18, отыскивая те клетки, которые могли активироваться бинокулярно. Ясно, что любая бинокулярная клетка этой области в правом полушарии должна получать входные сигналы как прямо от правого глаза (через НКТ), так и от левого глаза и левого полушария через мозолистое тело. Как выяснилось, рецептивное поле каждой бинокулярной клетки захватывало среднюю вертикаль сетчатки, причем та его часть, которая относится к левой половине поля зрения, доставляла информацию от правого глаза, а та, которая заходит в правую половину, — от левого глаза. Другие свойства клеток, исследованные в этом эксперименте, включая ориентационную избирательность, оказались идентичными (рис. 102). Полученные результаты ясно показали, что мозолистое тело связывает клетки друг с другом таким образом, чтобы их рецептивные поля могли заходить и вправо, и влево от средней вертикали. Таким образом, оно как бы склеивает две половины изображения окружающего мира. Чтобы лучше себе это представить, предположим, что изначально кора нашего мозга образовалась как одно целое, не разделенное на два полушария. В этом случае поле 17 имело бы вид одного непрерывного слоя, на который отображалось бы все зрительное поле. Тогда соседние клетки для реализации таких свойств как, например, чувствительность к движению и ориентационная избирательность, должны были бы иметь, разумеется, сложную систему взаимных связей. Теперь вообразим, что «конструктор» (будь то бог, или, скажем, естественный отбор) решил, что так дальше оставлять нельзя — отныне половина всех клеток должна образовать одно полушарие, а другая половина — другое полушарие. Что тогда нужно сделать со всем множеством межклеточных связей, если две совокупности клеток должны теперь отодвинуться друг от друга? По-видимому, можно просто растянуть эти связи, образовав из них часть мозолистого тела. Для того чтобы устранить задержку при передаче сигналов по такому длинному пути (у человека примерно 12–15 сантиметров), нужно увеличить скорость передачи, снабдив волокна миелиновой оболочкой. Разумеется, на самом деле ничего такого в процессе эволюции не происходило; задолго до того, как возникла кора, мозг уже имел два отдельных полушария. Эксперимент Берлукки и Риццолатти, на мой взгляд, дал одно из наиболее ярких подтверждений удивительной специфичности нейронных связей. Клетка, показанная на рис. 108 (около кончика электрода) и, вероятно, миллион других подобных клеток, соединенных через мозолистое тело, приобретают свою ориентационную избирательность как за счет локальных связей с соседними клетками, так и за счет связей, идущих через мозолистое тело из другого полушария от клеток с такой же ориентационной чувствительностью и сходным расположением рецептивных полей (сказанное относится и к другим свойствам клеток, таким как дирекциональная специфичность, способность реагировать на концы линий, а также сложность). Каждая из клеток зрительной коры, имеющих связи через мозолистое тело, должна получать входные сигналы от клеток другого полушария с точно такими же свойствами. Мы знаем множество фактов, указывающих на избирательность соединений в нервной системе, но я думаю, что данный пример — наиболее яркий и убедительный. Рассмотренные выше аксоны клеток зрительной коры составляют лишь небольшую долю всех волокон мозолистого тела. На соматосенсорной коре проводились эксперименты с использованием аксонного транспорта, аналогичные описанным в предыдущих главах опытам с инъекцией радиоактивной аминокислоты в глаз. Их результаты показывают, что мозолистое тело таким же образом связывает те участки коры, которые активируются кожными и суставными рецепторами, расположенными вблизи средней линии тела на туловище и голове, но не связывает корковые проекции конечностей. Каждая область коры соединяется с несколькими или даже многими другими областями коры того же полушария. Например, первичная зрительная кора связана с полем 18 (зрительной зоной 2), с медиальной височной областью (зоной MT), со зрительной зоной 4 и еще с одной или двумя областями. Многие участки коры имеют также связи с несколькими областями другого полушария, осуществляемые через мозолистое тело, а в некоторых случаях — через переднюю комиссуру. Поэтому мы можем рассматривать эти комиссуральные связи просто как особый вид кортико-кортикальных связей. Легко сообразить, что об этом свидетельствует такой простой пример: если я говорю вам, что моя левая рука ощущает холод или что я увидел что-то слева, то я формулирую слова, используя свои корковые речевые зоны, находящиеся в левом полушарии (сказанное, может быть, и не совсем верно, поскольку я левша); информация, поступающая от левой половины поля зрения или от левой руки, передается в мое правое полушарие; потом соответствующие сигналы должны быть переданы через мозолистое тело в речевую зону коры другого полушария, чтобы я мог сказать что-нибудь о своих ощущениях. В серии работ, начатых в начале 1960-х годов, Р. Сперри (сейчас он работает в Калифорнийском технологическом институте) и его сотрудники показали, что человек с перерезанным мозолистым телом (для лечения эпилепсии) теряет способность рассказывать о тех событиях, информация о которых попадает в правое полушарие. Работа с такими испытуемыми стала ценным источником новых сведений о различных функциях коры, включая мышление и сознание. Первые статьи об этом появились в журнале Brain; они чрезвычайно интересны, и их сможет без труда понять всякий, кто прочел настоящую книгу.
Стереоскопическое зрение
Механизм оценки удаленности, основанный на сравнении двух сетчаточных изображений, настолько надежен, что многие люди (если они не психологи и не специалисты по физиологии зрения) даже не подозревают о его существовании. Для того чтобы убедиться в важности этого механизма, попробуйте в течение нескольких минут вести автомобиль или велосипед, играть в теннис или прокатиться на лыжах, закрыв один глаз. Стереоскопы вышли из моды, и вы можете найти их только в антикварных магазинах. Однако большинство читателей смотрели стереоскопические фильмы (когда зрителю приходится надевать специальные очки). Принцип действия как стереоскопа, так и стереоскопических очков основан на использовании механизма стереопсиса. Изображения на сетчатках двумерны, а между тем мы видим мир трехмерным. Очевидно, что как для человека, так и для животных важна способность определять расстояние до объектов. Точно так же восприятие трехмерной формы предметов означает оценку относительной глубины. Рассмотрим в качестве простого примера круглый предмет. Если он расположен наклонно по отношению к линии взора, его изображение на сетчатках будет эллиптическим, однако обычно мы без труда воспринимаем такой предмет как круглый. Для этого необходима способность к восприятию глубины. Человек обладает многими механизмами оценки глубины. Некоторые из них столь очевидны, что вряд ли заслуживают упоминания. Тем не менее я их упомяну. Если приблизительно известна величина объекта, например в случае таких объектов, как человек, дерево или кошка, то можно оценить расстояние до него (правда, есть риск ошибиться, если мы столкнемся с карликом, карликовым деревом или львом). Если один предмет расположен впереди другого и частично его заслоняет, то мы воспринимаем передний объект как расположенный ближе. Если взять проекцию параллельных линий, например железнодорожных рельсов, уходящих вдаль, то в проекции они будут сближаться. Это пример перспективы — весьма эффективного показателя глубины. Выпуклый участок стены кажется более светлым в верхней своей части, если источник света расположен выше (обычно источники света и находятся вверху), а углубление в ее поверхности, если оно освещается сверху, кажется в верхней части более темным. Если же источник света поместить внизу, то выпуклость будет выглядеть как углубление, а углубление — как выпуклость. Важным признаком удаленности служит параллакс движения — кажущееся относительное смещение близких и более далеких предметов, если наблюдатель будет двигать головой влево и вправо или вверх и вниз. Если какой-то твердый предмет поворачивается, пусть даже на небольшой угол, то сразу же выявляется его трехмерная форма. Если мы фокусируем хрусталик нашего глаза на близко расположенном предмете, то более удаленный предмет будет не в фокусе; таким образом, меняя форму хрусталика, т.е. изменяя аккомодацию глаза (см. гл. 2 и 6), мы получаем возможность оценивать удаленность предметов. Если изменять относительное направление осей обоих глаз, сводя их или разводя (осуществляя конвергенцию или дивергенцию), то можно свести вместе два изображения предмета и удерживать их в этом положении. Таким образом, управляя либо хрусталиком, либо положением глаз, можно оценить удаленность объекта. На этих принципах основаны конструкции ряда дальномеров. За исключением конвергенции и дивергенции, все остальные показатели удаленности, перечисленные до сих пор, являются монокулярными. Наиболее важный механизм восприятия глубины — стереопсис — зависит от совместного использования двух глаз. При рассматривании любой трехмерной сцены два глаза формируют несколько различные изображения на сетчатке. Вы легко можете в этом убедиться, если будете смотреть прямо вперед и быстро перемещать голову из стороны в сторону примерно на 10 см или же быстро закрывать поочередно то один, то другой глаз. Если перед вами плоский объект, вы не заметите особой разницы. Однако, если сцена включает предметы на разном расстоянии от вас, вы заметите существенные изменения в картине. В процессе стереопсиса мозг сравнивает изображения одной и той же сцены на двух сетчатках и с большой точностью оценивает относительную глубину. Предположим, наблюдатель фиксирует взором некоторую точку P. Это утверждение эквивалентно тому, как если мы скажем: глаза направляются таким образом, чтобы изображения точки оказались в центральных ямках обоих глаз (F на рис. 103). Предположим теперь, что Q — это другая точка пространства, которая кажется наблюдателю расположенной на такой же глубине, что и P. Пусть QL и QR — изображения точки Q на сетчатках левого и правого глаза. В этом случае точки QL и QR называют корреспондирующими точками двух сетчаток. Очевидно, что две точки, совпадающие с центральными ямками сетчаток, будут корреспондирующими. Из геометрических соображений ясно также, что точка Q', оцениваемая наблюдателем как расположенная ближе, чем Q, будет давать на сетчатках две проекции — Q'L и Q'R — в некорреспондирующих точках, расположенных дальше друг от друга, чем в том случае, если бы эти точки были корреспондирующими (эта ситуация изображена в правой части рисунка). Точно так же, если рассматривать точку, расположенную дальше от наблюдателя, то окажется, что ее проекции на сетчатках будут расположены ближе друг к другу, чем корреспондирующие точки. То, что сказано выше о корреспондирующих точках, — это частично определения, а частично утверждения, вытекающие из геометрических соображений. При рассмотрении этого вопроса учитывается также психофизиология восприятия, поскольку наблюдатель субъективно оценивает, дальше или ближе точки P расположен объект. Введем еще одно определение. Все точки, которые, подобно точке Q (и, конечно, точке P), воспринимаются как равноудаленные, лежат на гороптере — поверхности, проходящей через точки P и Q, форма которой отличается как от плоскости, так и от сферы и зависит от нашей способности оценивать удаленность, т.е. от нашего мозга. Расстояния от центральной ямки F до проекций точки Q (QL и QR) близки, но не равны. Если бы они всегда были равны, то линия пересечения гороптера с горизонтальной плоскостью представляла бы собой круг.
 Рис. 103. Слева: если наблюдатель смотрит на точку P, то два ее изображения (проекции) попадают на центральные ямки двух глаз (точки F). Q — точка, которая, по оценке наблюдателя, находится на таком же расстоянии от него, что и P. В этом случае говорят, что две проекции точки Q (QL и QR) попадают в корреспондирующие точки сетчаток. (Поверхность, составленную из всех точек Q, которые кажутся находящимися на одинаковом расстоянии от наблюдателя, таком же, как точка P, называют гороптером, проходящим через точку P). Справа: если точка Q' находится ближе к наблюдателю, чем Q, то ее проекции на сетчатках (Q'L и Q'R) будут отстоять друг от друга по горизонтали дальше, чем если бы они находились в корреспондирующих точках. Если бы точка Q' находилась дальше, то проекции Q'L и Q'R оказались бы сдвинутыми по горизонтали ближе друг к другу.
Рис. 103. Слева: если наблюдатель смотрит на точку P, то два ее изображения (проекции) попадают на центральные ямки двух глаз (точки F). Q — точка, которая, по оценке наблюдателя, находится на таком же расстоянии от него, что и P. В этом случае говорят, что две проекции точки Q (QL и QR) попадают в корреспондирующие точки сетчаток. (Поверхность, составленную из всех точек Q, которые кажутся находящимися на одинаковом расстоянии от наблюдателя, таком же, как точка P, называют гороптером, проходящим через точку P). Справа: если точка Q' находится ближе к наблюдателю, чем Q, то ее проекции на сетчатках (Q'L и Q'R) будут отстоять друг от друга по горизонтали дальше, чем если бы они находились в корреспондирующих точках. Если бы точка Q' находилась дальше, то проекции Q'L и Q'R оказались бы сдвинутыми по горизонтали ближе друг к другу.
Предположим теперь, что мы фиксируем взглядом некоторую точку в пространстве и что в этом пространстве расположены два точечных источника света, которые дают проекцию на каждой сетчатке в виде световой точки, причем эти точки — не корреспондирующие: расстояние между ними несколько больше, чем между корреспондирующими точками. Любое такое отклонение от положения корреспондирующих точек мы будем называть диспаратностью. Если это отклонение в горизонтальном направлении не превышает 2° (0,6 мм на сетчатке), а по вертикали не больше нескольких угловых минут, то мы будем зрительно воспринимать одиночную точку в пространстве, расположенную ближе, чем та, которую мы фиксируем. Если же расстояния между проекциями точки будут не больше, а меньше, чем между корреспондирующими точками, то данная точка будет казаться расположенной дальше, чем точка фиксации. Наконец, в том случае, если вертикальное отклонение будет превышать несколько угловых минут или же горизонтальное будет больше 2°, то мы увидим две отдельные точки, которые, возможно, покажутся расположенными дальше или ближе точки фиксации. Эти экспериментальные результаты иллюстрируют основной принцип стереовосприятия, впервые сформулированный в 1838 году сэром Ч. Уитстоном (который также изобрел прибор, известный в электротехнике как «мостик Уитстона»). Кажется почти невероятным, что до этого открытия ни один человек, по-видимому, не отдавал себе отчета в том, что наличие едва заметных различий в изображениях, проецируемых на сетчатки двух глаз, может приводить к отчетливому впечатлению глубины. Такой стереоэффект может продемонстрировать за несколько минут любой человек, способный произвольно сводить или разводить оси своих глаз, или же тот, у кого есть карандаш, кусок бумаги и несколько небольших зеркал или призм. Непонятно, как прошли мимо этого открытия Евклид, Архимед и Ньютон. В своей статье Уитстон отмечает, что Леонардо да Винчи был очень близок к открытию этого принципа. Леонардо указывал, что шар, расположенный перед какой-либо пространственной сценой, виден каждым глазом по-разному — левым глазом мы немного дальше видим его левую сторону, а правым глазом — правую. Далее Уитстон отмечает, что если бы вместо шара Леонардо выбрал куб, то он, безусловно, заметил бы, что его проекции для разных глаз различны. После этого он мог бы, как и Уитстон, заинтересоваться тем, что будет, если специально спроецировать два подобных изображения на сетчатки двух глаз. Важным физиологическим фактом является то, что ощущение глубины (т.е. возможность «непосредственно» видеть, дальше или ближе точки фиксации расположен тот или иной объект) возникает в тех случаях, когда два сетчаточных изображения несколько смещены относительно друг друга в горизонтальном направлении — раздвинуты или, наоборот, сближены (если только это смещение не превышает примерно 2°, а вертикальное смещение близко к нулю). Это, разумеется, соответствует геометрическим соотношениям: если по отношению к некоторой точке отсчета расстояния объект расположен ближе или дальше, то его проекции на сетчатках будут раздвинуты или сближены по горизонтали, тогда как существенного вертикального смещения изображений не произойдет. На этом и основано действие стереоскопа, изобретенного Уитстоном. Стереоскоп в течение примерно полувека был настолько популярен, что имелся чуть ли не в каждом доме. Тот же принцип лежит в основе и стереокино, которое мы сейчас смотрим, используя для этого специальные поляроидные очки. В первоначальной конструкции стереоскопа наблюдатель рассматривал два изображения, помещенные в ящик, с помощью двух зеркал, которые были расположены таким образом, что каждый глаз видел только одно изображение. Для удобства теперь часто используют призмы и фокусирующие линзы. Два изображения идентичны во всем, кроме небольших горизонтальных смещений, которые и создают впечатление глубины. Любой может изготовить фотографию, пригодную для использования в стереоскопе, если выберет какой-либо неподвижный объект (или сцену), сделает снимок, а затем сдвинет фотоаппарат на 5 сантиметров вправо или влево и сделает второй снимок. Не все обладают способностью воспринимать глубину с помощью стереоскопа. Вы сами можете легко проверить свой стереопсис, если воспользуетесь стереопарами, приведенными на рис. 105 и 106. Если у вас есть стереоскоп, вы можете сделать копии изображенных здесь стереопар и вставить их в стереоскоп. Вы можете также поместить тонкий кусок картона перпендикулярно между двумя изображениями из одной стереопары и попытаться смотреть каждым глазом на свое изображение, установив глаза параллельно, как если бы вы смотрели вдаль. Можно также научиться сводить и разводить глаза с помощью пальца, поместив его между глазами и стереопарой и передвигая вперед или назад, пока изображения не сольются, после чего (это самое трудное) вы сможете рассматривать слитое изображение, стараясь, чтобы оно не разделилось на два. Если у вас это получится, то кажущиеся отношения глубины будут противоположны тем, которые воспринимаются при использовании стереоскопа.

 Рис. 104. А. Стереоскоп Уитстона. Б. Схема стереоскопа Уитстона, составленная им самим. Наблюдатель сидит перед двумя зеркалами (А и А'), поставленными под углом 40° к направлению его взора, и смотрит на две совмещенные в поле зрения картинки — Е (правым глазом) и Е' (левым глазом). В созданном позже более простом варианте две картинки помещаются рядом так, что расстояние между их центрами примерно равно расстоянию между глазами. Две призмы отклоняют направление взора так, что при надлежащей конвергенции левый глаз видит левое изображение, а правый глаз — правое изображение. Вы сами можете попробовать обойтись без стереоскопа, представив себе, что смотрите на очень удаленный предмет глазами, оси которых установлены параллельно друг другу. Тогда левый глаз будет смотреть на левое изображение, а правый — на правое.
Рис. 104. А. Стереоскоп Уитстона. Б. Схема стереоскопа Уитстона, составленная им самим. Наблюдатель сидит перед двумя зеркалами (А и А'), поставленными под углом 40° к направлению его взора, и смотрит на две совмещенные в поле зрения картинки — Е (правым глазом) и Е' (левым глазом). В созданном позже более простом варианте две картинки помещаются рядом так, что расстояние между их центрами примерно равно расстоянию между глазами. Две призмы отклоняют направление взора так, что при надлежащей конвергенции левый глаз видит левое изображение, а правый глаз — правое изображение. Вы сами можете попробовать обойтись без стереоскопа, представив себе, что смотрите на очень удаленный предмет глазами, оси которых установлены параллельно друг другу. Тогда левый глаз будет смотреть на левое изображение, а правый — на правое.
Даже если вам не удастся повторить опыт с восприятием глубины — из-за того ли, что у вас нет стереоскопа, или потому, что вы не можете произвольно сводить и разводить оси глаз, — вы все-таки сможете понять суть дела, хотя не получите удовольствия от стереоэффекта. В верхней стереопаре на рис. 105 в двух квадратных рамках имеется по небольшому кружку, один из которых смещен немного влево от центра, а другой — немного вправо. Если рассматривать эту стереопару двумя глазами, используя стереоскоп или иной метод совмещения изображений, то вы увидите кружок не в плоскости листа, а впереди него на расстоянии около 2,5 см. Если так же рассматривать нижнюю стереопару на рис. 105, то кружок будет виден позади плоскости листа. Вы воспринимаете положение кружка таким образом потому, что на сетчатки ваших глаз попадает в точности такая же информация, как если бы кружок действительно находился впереди или позади плоскости рамки.
 Рис. 105. Если верхнюю стереопару вставить в стереоскоп, то кружок будет выглядеть расположенным впереди плоскости рамки. В нижней стереопаре он будет располагаться позади плоскости рамки. (Вы можете проделать такой опыт без стереоскопа, путем конвергенции или дивергенции глаз; для большинства людей конвергенция легче. Для облегчения задачи можно взять кусок картона и поставить его между двумя изображениями стереопары. Поначалу это упражнение может показаться вам трудным и утомительным; не усердствуйте при первой попытке. При конвергенции глаз на верхней стереопаре кружочек будет виден дальше плоскости, а на нижней — ближе).
Рис. 105. Если верхнюю стереопару вставить в стереоскоп, то кружок будет выглядеть расположенным впереди плоскости рамки. В нижней стереопаре он будет располагаться позади плоскости рамки. (Вы можете проделать такой опыт без стереоскопа, путем конвергенции или дивергенции глаз; для большинства людей конвергенция легче. Для облегчения задачи можно взять кусок картона и поставить его между двумя изображениями стереопары. Поначалу это упражнение может показаться вам трудным и утомительным; не усердствуйте при первой попытке. При конвергенции глаз на верхней стереопаре кружочек будет виден дальше плоскости, а на нижней — ближе).
В 1960 году Бела Юлеш из фирмы Bell Telephone Laboratories придумал весьма полезную и изящную методику для демонстрации стереоэффекта. Изображение, представленное на рис. 107, на первый взгляд кажется однородной случайной мозаикой из маленьких треугольничков. Так оно и есть, за исключением того, что в центральной части имеется скрытый треугольник большего размера. Если вы будете рассматривать это изображение с помощью двух кусочков цветного целлофана, помещенных перед глазами, — красного перед одним глазом и зеленого перед другим, то вы должны увидеть в центре треугольник, выступающий из плоскости листа вперед, как в предыдущем случае с маленьким кружком на стереопарах. (Быть может, в первый раз вам придется смотреть минуту или около этого, пока не возникнет стереоэффект.) Если поменять куски целлофана местами, произойдет инверсия глубины. Ценность этих стереопар Юлеша заключается в том, что если у вас нарушено стереовосприятие, то вы не увидите треугольника впереди или позади окружающего фона.
 Рис. 106. Еще одна стереопара.
Рис. 106. Еще одна стереопара.
Подводя итоги, можно сказать, что наша способность ощущать стереоэффект зависит от пяти условий: 1. Имеется много косвенных признаков глубины — частичное заслонение одних предметов другими, параллакс движения, вращение предмета, относительные размеры, отбрасывание теней, перспектива. Однако наиболее мощным механизмом является стереопсис. 2. Если мы фиксируем взглядом какую-то точку в пространстве, то проекции этой точки попадают в центральные ямки обеих сетчаток. Любая точка, которая оценивается как расположенная на том же расстоянии от глаз, что и точка фиксации, образует две проекции в корреспондирующих точках сетчаток. 3. Стереоэффект определяется простым геометрическим фактом — если некоторый объект находится ближе точки фиксации, то две его проекции на сетчатках оказываются дальше друг от друга, чем корреспондирующие точки. 4. Главный вывод, основанный на результатах экспериментов с испытуемыми, заключается в следующем: объект, проекции которого на сетчатках правого и левого глаза попадают на корреспондирующие точки, воспринимается как расположенный на том же расстоянии от глаз, что и точка фиксации; если проекции этого объекта раздвинуты по сравнению с корреспондирующими точками, объект кажется расположенным ближе точки фиксации; если же они, наоборот, сближены, объект кажется расположенным дальше точки фиксации. 5. При горизонтальном смещении проекций больше чем на 2° или вертикальном смещении больше нескольких угловых минут возникает двоение.
 Рис. 107. Для того чтобы получить это изображение, называемое анаглифом, Бела Юлес сначала построил две системы случайно расположенных маленьких треугольников; они различались только тем, что 1) в одной системе были красные треугольники на белом фоне, а в другой — зеленые на белом фоне; 2) в пределах большой треугольной зоны (вблизи центра рисунка) все зеленые треугольники несколько смещены влево по сравнению с красными. После этого две системы совмещаются, но с небольшим сдвигом, так что сами треугольники не накладываются друг на друга. Если на полученное изображение смотреть через зеленый фильтр из целлофана, то будут видны только красные элементы, а если через красный фильтр — только зеленые. Если же перед одним глазом поместить зеленый фильтр, а перед другим — красный, то вы увидите большой треугольник, выступающий примерно на 1 см перед страницей. В случае перемены фильтров местами треугольник будет виден за плоскостью страницы.
Рис. 107. Для того чтобы получить это изображение, называемое анаглифом, Бела Юлес сначала построил две системы случайно расположенных маленьких треугольников; они различались только тем, что 1) в одной системе были красные треугольники на белом фоне, а в другой — зеленые на белом фоне; 2) в пределах большой треугольной зоны (вблизи центра рисунка) все зеленые треугольники несколько смещены влево по сравнению с красными. После этого две системы совмещаются, но с небольшим сдвигом, так что сами треугольники не накладываются друг на друга. Если на полученное изображение смотреть через зеленый фильтр из целлофана, то будут видны только красные элементы, а если через красный фильтр — только зеленые. Если же перед одним глазом поместить зеленый фильтр, а перед другим — красный, то вы увидите большой треугольник, выступающий примерно на 1 см перед страницей. В случае перемены фильтров местами треугольник будет виден за плоскостью страницы.
Физиология стереоскопического зрения
Если мы хотим знать, каковы мозговые механизмы стереопсиса, то проще всего начать с вопроса: существуют ли нейроны, реакции которых специфически определяются относительным горизонтальным смещением изображений на сетчатках двух глаз? Посмотрим сначала, как отвечают клетки нижних уровней зрительной системы при одновременной стимуляции обоих глаз. Мы должны начать с нейронов поля 17 или более высокого уровня, поскольку ганглиозные клетки сетчатки явно монокулярные, а клетки наружного коленчатого тела, в котором входы от правого и левого глаз распределены по разным слоям, тоже можно считать монокулярными — они отвечают на стимуляцию либо одного глаза, либо другого, но не обоих одновременно. В поле 17 примерно половину нейронов составляют бинокулярные клетки, отвечающие на стимуляцию обоих глаз. При тщательном тестировании выясняется, что ответы этих клеток, по-видимому, мало зависят от относительного положения проекций стимулов на сетчатках двух глаз. Рассмотрим типичную сложную клетку, которая отвечает непрерывным разрядом на движение стимульной полосы через ее рецептивное поле в том или другом глазу. При одновременной стимуляции обоих глаз частота разрядов этой клетки выше, чем при стимуляции одного глаза, но обычно для ответа такой клетки несущественно, попадают ли в какой-то момент проекции стимула точно в одни и те же участки двух рецептивных полей. Наилучший ответ регистрируется тогда, когда эти проекции входят и выходят из соответствующих рецептивных полей двух глаз примерно в одно время; однако не столь важно, которая из проекций немного опережает другую. На рис. 108 показана характерная кривая зависимости ответа (например, общего числа импульсов в ответе за одно прохождение стимула через рецептивное поле) от разницы в положении стимула на обеих сетчатках. Эта кривая очень близка к горизонтальной прямой, из чего ясно, что относительное положение стимулов на двух сетчатках не очень существенно. Клетка такого типа будет хорошо реагировать на линию надлежащей ориентации независимо от ее удаленности — расстояние до линии может быть больше, равно или меньше расстояния до точки, фиксируемой взором.
 Рис. 108. При одновременной стимуляции обоих глаз вертикальной светлой линией, движущейся влево, обычная бинокулярная клетка в поле 17 будет давать одинаковые ответы при трех разных относительных положениях этой линии на обеих сетчатках. Нулевая диспаратность означает, что различия в положении нет, — как если бы обезьяна смотрела на экран, на котором предъявляются стимулы. Отсутствие диспаратности не приводит к заметному усилению ответа клетки.
Рис. 108. При одновременной стимуляции обоих глаз вертикальной светлой линией, движущейся влево, обычная бинокулярная клетка в поле 17 будет давать одинаковые ответы при трех разных относительных положениях этой линии на обеих сетчатках. Нулевая диспаратность означает, что различия в положении нет, — как если бы обезьяна смотрела на экран, на котором предъявляются стимулы. Отсутствие диспаратности не приводит к заметному усилению ответа клетки.
По сравнению с этой клеткой нейроны, ответы которых представлены на рис. 109 и 110, весьма чувствительны к относительному положению двух стимулов на двух сетчатках, т.е. чувствительны к глубине. Первый нейрон (рис. 109) лучше всего отвечает в том случае, если стимулы попадают точно на корреспондирующие участки двух сетчаток. Величина несовмещения стимулов по горизонтали (т.е. диспаратность), при которой клетка уже перестает реагировать, составляет некоторую долю ширины ее рецептивного поля. Поэтому клетка отвечает тогда и только тогда, когда объект находится примерно на таком же расстоянии от глаз, как и точка фиксации. Второй нейрон (рис. 110) отвечает только тогда, когда объект расположен дальше точки фиксации. Имеются также клетки, отвечающие только тогда, когда стимул расположен ближе этой точки. При изменении степени диспаратности нейроны двух последних типов, называемые дальними клетками и ближними клетками, очень резко изменяют интенсивность своих ответов в точке нулевой диспаратности или поблизости он нее. Нейроны всех трех типов (клетки, настроенные на диспаратность) были обнаружены в поле 17 обезьяны. Пока не совсем ясно, насколько часто они там встречаются, расположены ли они в определенных слоях коры и находятся ли в определенных пространственных отношениях к колонкам глазодоминантности. Эти клетки весьма чувствительны к расстоянию объекта от глаз, которое кодируется в виде относительного положения соответствующих стимулов на двух сетчатках. Еще одна особенность этих клеток — то, что они не отвечают на стимуляцию только одного глаза или же отвечают, но очень слабо. Все эти клетки обладают общим свойством ориентационной избирательности; насколько нам известно, они сходны с обычными сложными клетками верхних слоев коры, но обладают еще дополнительным свойством — чувствительностью к глубине. Кроме того, эти клетки хорошо реагируют на движущиеся стимулы, а иногда и на концы линий.
 Рис. 109. В ответе этой избирательно отвечающей клетки наблюдается значительная разница в зависимости от того, находится ли стимул точно на том же расстоянии, что и точка фиксации, или же он расположен ближе или дальше. Данная клетка отвечает только тогда, когда расстояние до стимула такое же, как и до точки фиксации. В этом эксперименте ось одного глаза отклоняли в горизонтальном направлении с помощью призмы; однако к такому же результату привело бы и смещение экрана по направлению к животному или от него.
Рис. 109. В ответе этой избирательно отвечающей клетки наблюдается значительная разница в зависимости от того, находится ли стимул точно на том же расстоянии, что и точка фиксации, или же он расположен ближе или дальше. Данная клетка отвечает только тогда, когда расстояние до стимула такое же, как и до точки фиксации. В этом эксперименте ось одного глаза отклоняли в горизонтальном направлении с помощью призмы; однако к такому же результату привело бы и смещение экрана по направлению к животному или от него.
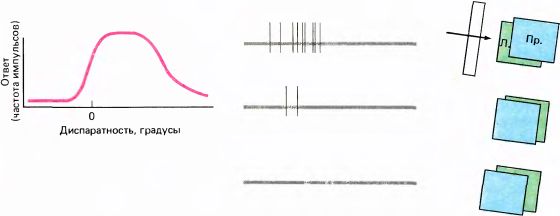 Рис. 110. У клетки представленного здесь типа («дальней») объекты, расположенные ближе плоскости экрана, вызывают очень слабый ответ или вовсе его не вызывают. При нулевой диспаратности (т.е. при расстоянии до стимула, равном расстоянию до экрана) небольшое смещение экрана сильно влияет на реакцию: она резко возрастает и остается постоянной для стимулов, расположенных дальше точки фиксации. Если отвести стимул достаточно далеко, то два рецептивных поля клетки уже не перекрываются, т.е., по существу, два глаза стимулируются независимо. Ответ в этом случае прекращается.
Рис. 110. У клетки представленного здесь типа («дальней») объекты, расположенные ближе плоскости экрана, вызывают очень слабый ответ или вовсе его не вызывают. При нулевой диспаратности (т.е. при расстоянии до стимула, равном расстоянию до экрана) небольшое смещение экрана сильно влияет на реакцию: она резко возрастает и остается постоянной для стимулов, расположенных дальше точки фиксации. Если отвести стимул достаточно далеко, то два рецептивных поля клетки уже не перекрываются, т.е., по существу, два глаза стимулируются независимо. Ответ в этом случае прекращается.
Дж. Поджо из медицинской школы Джонса Гопкинса регистрировал ответы таких клеток в поле 17 бодрствующей обезьяны с вживленными электродами, которая была предварительно обучена фиксировать взглядом определенный объект. У наркотизированных обезьян такие клетки тоже выявлялись в коре, но редко встречались в поле 17 и очень часто — в поле 18. Я был бы крайне удивлен, если бы оказалось, что животные и человек могут стереоскопически оценивать расстояния до объектов с помощью только трех описанных выше типов клеток — настроенных на нулевую диспаратность, «ближних» и «дальних». Я бы скорее ожидал найти полный набор клеток для всех возможных глубин. У бодрствующих обезьян Поджо встречал также узконастроенные клетки, которые лучше всего реагировали не на нулевую диспаратность, а на небольшие отклонения от нее; по-видимому, в коре могут быть специфические нейроны для всех ступеней диспаратности. Хотя мы до сих пор не знаем, как именно мозг «реконструирует» сцену, включающую множество разноудаленных объектов (что бы мы ни понимали под словом «реконструкция»), клетки вроде описанных выше, вероятно, участвуют в первых этапах этого процесса.
Некоторые проблемы, связанные со стереоскопическим зрением
За время изучения стереопсиса психофизики столкнулись с целым рядом проблем. Оказалось, что обработка некоторых бинокулярных стимулов происходит в зрительной системе совершенно непонятными способами. Я мог бы привести много примеров такого рода, но ограничусь лишь двумя. На примере стереопар, приведенных на рис. 105, мы видели, что смещение двух идентичных изображений (в данном случае кружков) по направлению друг к другу приводит к ощущению большей близости, а по направлению друг от друга — к ощущению большей удаленности. Предположим теперь, что мы делаем одновременно обе эти операции, для чего помещаем в каждую рамку по два кружка, расположенных друг около друга (рис. 111). Очевидно, что рассматривание такой стереопары могло бы привести к восприятию двух кружков — одного ближе, а другого дальше плоскости фиксации. Однако можно предположить и другой вариант: мы увидим просто два кружка, лежащие рядом в плоскости фиксации. Дело в том, что этим двум пространственным ситуациям соответствуют одинаковые изображения на сетчатках. В действительности данная пара стимулов может быть воспринята только как два кружка в плоскости фиксации, в чем легко убедиться, если любым способом достичь слияния квадратных рамок на рис. 111. Точно так же можно представить себе ситуацию, когда мы рассматриваем две цепочки из знаков ×, скажем, по шесть знаков в цепочке. Если рассматривать их в стереоскоп, то в принципе можно воспринять любую из ряда возможных конфигураций в зависимости от того, какой знак × из левой цепочки сольется с определенным знаком × в правой цепочке. На самом же деле, если мы будем рассматривать такую стереопару в стереоскоп (или иным способом, создающим стереоэффект), то всегда увидим шесть знаков × в плоскости фиксации. Мы до сих пор не знаем, как мозг разрешает эту неоднозначность и выбирает простейшую из всех возможных комбинаций. Из-за такого рода неоднозначностей трудно даже вообразить, как нам удается воспринимать объемной сцену, включающую множество ветвей разной величины, находящихся на разных расстояниях от нас. Правда, физиологические данные подсказывают, что задача, возможно, не столь трудна, так как разные ветви скорее всего будут иметь разную ориентацию, а мы уже знаем, что клетки, участвующие в стереопсисе, всегда бывают ориентационно-избирательными.
 Рис. 111. Стереопары, приведенные на рис. 105, вызывали ощущение, что кружок расположен ближе или дальше плоскости рамки. Здесь эти две стереопары объединены, и поэтому, казалось бы, мы должны видеть один кружок ближе, а другойдальше. Однако на самом деле этого не получается; оба кружка видны на том же расстоянии, что и рамка.
Рис. 111. Стереопары, приведенные на рис. 105, вызывали ощущение, что кружок расположен ближе или дальше плоскости рамки. Здесь эти две стереопары объединены, и поэтому, казалось бы, мы должны видеть один кружок ближе, а другойдальше. Однако на самом деле этого не получается; оба кружка видны на том же расстоянии, что и рамка.
 Рис. 112. Эту стереопару невозможно слить в единое восприятие, как другие стереопары (например, показанные на рис. 105 и 106). Вместо этого получается эффект «борьбы полей зрения» — вы видите мозаичную картину, состоящую из фрагментов обоих изображений и беспрестанно меняющуюся.
Рис. 112. Эту стереопару невозможно слить в единое восприятие, как другие стереопары (например, показанные на рис. 105 и 106). Вместо этого получается эффект «борьбы полей зрения» — вы видите мозаичную картину, состоящую из фрагментов обоих изображений и беспрестанно меняющуюся.
Второй пример непредсказуемости бинокулярных эффектов, имеющий отношение к стереопсису, — это так называемая борьба полей зрения, о которой мы упоминаем также в разделе о косоглазии (гл. 9). Если на сетчатках правого и левого глаза создаются очень сильно различающиеся изображения, то часто одно из них перестает восприниматься. Если вы будете смотреть левым глазом на решетку из вертикальных линий, а правым глазом — на решетку из горизонтальных линий (рис. 112; можно пользоваться стереоскопом или конвергенцией глаз), то, казалось бы, следует ожидать, что вы увидите сетку из пересекающихся линий. Однако в действительности почти невозможно увидеть оба набора линий одновременно. Виден или тот или другой, причем каждый из них — лишь в течение нескольких секунд, после чего он исчезает и появляется другой. Иногда можно также увидеть как бы мозаику из двух этих изображений, в которой отдельные однородные участки будут перемещаться, сливаться или разделяться, а ориентация линий в них будет меняться (см. рис. 112, внизу). По какой-то причине нервная система не может воспринимать столь разные стимулы одновременно в одном и том же участке поля зрения, и она подавляет переработку одного из них. Слово «подавлять» мы используем здесь просто как иное описание того же феномена: на самом деле мы не знаем, как осуществляется такое подавление и на каком уровне центральной нервной системы оно происходит. Мне думается, мозаичный характер воспринимаемого образа при борьбе полей зрения позволяет предположить, что «принятие решений» в этом процессе происходит на достаточно ранних этапах переработки зрительной информации, возможно, в поле 17 или 18. (Я рад, что мне не нужно защищать это предположение.) Феномен борьбы полей зрения означает, что в тех случаях, когда зрительная система не может объединить изображения на двух сетчатках (в плоскую картину, если изображения одинаковы, или в трехмерную сцену, если имеется лишь небольшая горизонтальная диспаратность), она просто отвергает один из образов — либо полностью, когда, например, мы смотрим в микроскоп, держа второй глаз открытым, либо частично или на время, как в примере, описанном выше. В ситуации с микроскопом существенную роль играет внимание, но нейронные механизмы, лежащие в основе такого переключения внимания, тоже неизвестны. Еще один пример борьбы полей зрения вы можете наблюдать, если просто будете рассматривать какую-нибудь многоцветную сцену или картину через очки с красным и зеленым светофильтрами. Впечатления разных наблюдателей в этом случае могут быть весьма различными, однако большинство людей (в том числе и я) отмечает переходы от общего красноватого тона к зеленоватому и обратно, но без желтого цвета, который получается при обычном смешении красного света с зеленым (см. гл. 8, рис. 121).
Стереослепота
Если человек слеп на один глаз, то очевидно, что он не будет обладать стереоскопическим зрением. Однако его нет и у некоторой части людей, зрение которых в остальных отношениях нормально. Удивительно то, что доля таких людей не слишком мала. Так, если показать стереопары вроде тех, что приводились на рис. 105 и 106, сотне студентов-испытуемых (применяя поляроиды и поляризованный свет), то обычно оказывается, что четыре или пять из них не могут достичь стереоэффекта. Нередко это удивляет их самих, так как в повседневных условиях они не испытывают никаких неудобств. Последнее может показаться странным любому, кто ради эксперимента пытался управлять автомобилем, закрыв один глаз. По-видимому, отсутствие стереопсиса достаточно хорошо компенсируется использованием других признаков глубины, таких как параллакс движения, перспектива, частичное закрытие одних предметов другими и т.п. В главе 9 мы рассмотрим случаи врожденного косоглазия, когда глаза длительное время работают несогласованно. Это может приводить к нарушению связей в коре, обеспечивающих бинокулярное взаимодействие, и в результате — к утрате стереопсиса. Косоглазие встречается не так уж редко, и даже слабая его степень, которая может остаться незамеченной, в некоторых случаях, вероятно, бывает причиной стереослепоты. В других же случаях нарушение стереопсиса, подобно цветовой слепоте, может быть наследственным. Поскольку в этой главе шла речь и о мозолистом теле, и о стереоскопическом зрении, я воспользуюсь случаем сказать кое-что о связи этих двух вещей. Попробуйте задать себе вопрос: каких нарушений стереопсиса можно ожидать у человека с перерезанным мозолистым телом? Ответ на этот вопрос ясен из схемы, приведенной на рис. 113.
 Рис. 113. Перерезка мозолистого тела приводит к потере стереопсиса в закрашенной части видимого пространства.
Рис. 113. Перерезка мозолистого тела приводит к потере стереопсиса в закрашенной части видимого пространства.
 Рис. 114. Результаты продольной перерезки хиазмы по средней линии. Испытуемый совсем не будет видеть две более темные зоны по краям рисунка, слева и справа. Между этими зонами, там, где окраска светлее, не будет стереопсиса, за исключением небольшой зоны треугольной формы за точкой P (здесь вообще ничего не видно) и зоны впереди точки P (здесь стереопсис сохранится).
Рис. 114. Результаты продольной перерезки хиазмы по средней линии. Испытуемый совсем не будет видеть две более темные зоны по краям рисунка, слева и справа. Между этими зонами, там, где окраска светлее, не будет стереопсиса, за исключением небольшой зоны треугольной формы за точкой P (здесь вообще ничего не видно) и зоны впереди точки P (здесь стереопсис сохранится).
Если человек фиксирует взглядом точку P, то проекции точки Q, расположенной ближе к глазам в пределах острого угла FPF, — QL и QR — окажутся в левом и в правом глазу по разные стороны от центральной ямки. Соответственно проекция QL передает информацию в левое полушарие, а проекция QR — в правое полушарие. Для того чтобы увидеть, что точка Q ближе, чем P (т.е. получить стереоэффект), нужно объединить информацию левого и правого полушарий. Но единственный способ сделать это — передать информацию по мозолистому телу. Если же путь через мозолистое тело разрушен, человек окажется стереослепым в закрашенной на рисунке области. В 1970 году Д. Митчелл и К. Блейкмор из Калифорнийского университета в Беркли исследовали стереоскопическое зрение у одного человека с перерезанным мозолистым телом и получили в точности предсказанный выше результат. Второй вопрос, тесно связанный с первым, состоит в том, какое нарушение стереопсиса произойдет, если перерезать по средней линии зрительную хиазму (что проделал Р. Майерс на кошках). Результат здесь будет в определенном смысле противоположным. Из рис. 114 должно быть ясно, что в этом случае каждый глаз станет слепым в отношении стимулов, падающих на носовую область сетчатки, т.е. исходящих из височной части поля зрения. Поэтому стереопсиса не будет в области пространства, окрашенной светлее, где он в норме имеется. Боковые зоны за пределами этой области вообще доступны только для одного глаза, так что стереопсис здесь отсутствует и в нормальных условиях, а после перерезки хиазмы они будут зонами слепоты (на рисунке это показано более темным цветом). В области позади точки фиксации, где перекрываются височные части полей зрения, ставшие теперь невидимыми, тоже наступит слепота. Однако в зоне ближе точки фиксации сохранившиеся полуполя обоих глаз перекрываются, так что здесь должен сохраниться стереопсис, если только не повреждено мозолистое тело. К. Блейкмор нашел все-таки больного с полной перерезкой хиазмы по средней линии (этот больной, будучи ребенком, получил перелом черепа при езде на велосипеде, что, по-видимому, привело к продольному разрыву хиазмы). При проверке у него была обнаружена именно та комбинация дефектов зрения, которую мы только что гипотетически описали.
8. Цветовое зрение
Сотни дополнительных долларов, которые покупатели соглашаются платить за цветной телевизор, предпочитая его черно-белому, означают, что цветовые ощущения для нас достаточно важны. Сложный аппарат глаза и мозга может воспринимать различия в спектральном составе света, отражаемого от видимых предметов, и легко представить себе, какие преимущества давала эта способность нашим предкам. Одним из преимуществ, несомненно, было то, что она затрудняла маскировку другим животным: потенциальной добыче намного труднее слиться с окружающим фоном, если хищник может различать не только интенсивность света, но и цвет. Столь же важным цвет может быть при поиске растительной пищи: обезьяна легко найдет ярко-красную ягоду, выделяющуюся среди зеленой листвы, и это даст животному несомненное преимущество, как, впрочем, и растению, поскольку семена проходят невредимыми через пищеварительный тракт обезьяны и рассеиваются на обширной площади. Для некоторых животных цвет важен при размножении; примерами служат ярко-красная окраска области промежности у макаков и изумительное оперение у самцов многих птиц. У людей давление отбора, направленное на сохранение или улучшение цветового зрения, видимо, ослабевает, судя по тому, что 7 или 8 процентов мужчин частично или полностью лишены цветового зрения, но отлично без него обходятся, причем этот дефект часто долгие годы остается незамеченным и выявляется лишь после того, как за рулем они проедут на красный свет. Даже те из нас, кто обладает нормальным цветовым зрением, могут испытывать подлинное наслаждение от черно-белых фильмов, которые в художественном отношении иногда могут быть шедеврами киноискусства. Как мы увидим позже, при слабом освещении все мы в цветовом отношении слепы. Ощущение цвета у позвоночных встречается спорадически; вероятно, в ходе эволюции оно неоднократно редуцировалось или даже исчезало, чтобы потом появиться снова. К млекопитающим, у которых цветовое зрение слабо развито или отсутствует, относятся мыши, крысы, кролики, кошки, собаки и ночная обезьяна дурукули. У сусликов и приматов, включая людей, человекообразных и большинство других обезьян, цветовое зрение хорошо развито. Из ночных животных, зрение которых приспособлено к слабому свету, лишь немногие хорошо различают цвета; это позволяет думать, что по каким-то причинам различение цветов и способность видеть при слабом свете несовместимы друг с другом. Среди других позвоночных цветовое зрение хорошо развито у многих рыб и птиц, но, вероятно, отсутствует или слабо выражено у рептилий и амфибий. Цветовым зрением обладают многие насекомые, в том числе мухи и пчелы. В отношении подавляющего большинства животных у нас нет точных сведений о способности различать цвета — вероятно, потому, что проводить поведенческие или физиологические тесты на цветовое зрение не так легко. Рис. 115. Цвет используется в живой природе для разных целей, и некоторые из них пока не известны. Синие пятнышки на боках этой рыбы (Hypsypops) становятся все менее яркими, по мере того как рыба растет, и исчезают, когда она достигает зрелости. Какое значение имеют эти пятнышки для других особей того же вида, неизвестно.
Рис. 115. Цвет используется в живой природе для разных целей, и некоторые из них пока не известны. Синие пятнышки на боках этой рыбы (Hypsypops) становятся все менее яркими, по мере того как рыба растет, и исчезают, когда она достигает зрелости. Какое значение имеют эти пятнышки для других особей того же вида, неизвестно.
Вопросом о цветовом зрении — несоразмерно его биологическому значению для человека — занимался ряд блестящих умов, включая Ньютона, Гёте (сильной стороной которого не были, однако, естественные науки) и Гельмгольца. Тем не менее до сих пор даже художники, физики и биологи часто плохо представляют себе, что такое цвет. Проблема возникает еще в детстве, когда нам впервые дают коробку с красками, а затем говорят, что желтый, синий и красный — это «основные» цвета и что желтый с синим дают зеленый. Многие из нас впоследствии поражаются кажущемуся противоречию с этим фактом, когда с помощью пары проекторов мы отбрасываем на экран два перекрывающихся пятна, желтое и синее, и видим в области их наложения красивый белый цвет. Результат смешения красок — это предмет физики; смешение же световых лучей — в основном вопрос биологии. Рассуждая о цвете, полезно мысленно разделять эти два аспекта — физический и биологический. Физика, которую нам при этом следует знать, ограничивается лишь некоторыми фактами о световых волнах. Биология же включает психофизику и физиологию. Психофизику интересуют наши чувства как детекторы внешней информации, а физиологию — лежащие в их основе внутренние механизмы, в частности работа нашей зрительной системы. Мы многое знаем о физике и психофизике цвета, но физиология находится все еще на сравнительно примитивном уровне, главным образом из-за того, что необходимые методы стали доступны лишь в последние десятилетия.
Природа света
Свет состоит из частиц, называемых фотонами, каждую из которых можно рассматривать как пакет электромагнитных волн. Будет ли луч электромагнитной энергии именно светом, а не рентгеновскими лучами или радиоволнами, определяется длиной волны — расстоянием от одного гребня волны до следующего: в случае света это расстояние составляет приблизительно 0,0000001 (10–7) метра, или 0,0005 миллиметра, или 0,5 микрометра, или 500 нанометров (нм). Свет — это по определению то, что мы можем видеть. Наши глаза могут воспринимать электромагнитные волны длиной от 400 до 700 нм. Обычно попадающий в наши глаза свет состоит из сравнительно однородной смеси лучей с различными длинами волн; такую смесь называют белым светом (хотя это весьма нестрогое понятие). Для оценки волнового состава световых лучей измеряют световую энергию, заключенную в каждом из последовательных небольших интервалов, например от 400 до 410 нм, от 410 до 420 нм и т.д., после чего рисуют график распределения энергии по длинам волн. Для света, приходящего от Солнца, этот график похож на левую кривую на рис. 116. Это кривая без резких подъемов и спадов с пологим максимумом в области 600 нм. Такая кривая типична для излучения раскаленного объекта. Положение максимума зависит от температуры источника: для Солнца это будет область около 600 нм, а для звезды более горячей, чем наше Солнце, максимум сдвинется к более коротким волнам — к голубому концу спектра, т.е. на нашем графике — влево. (Представление художников о том, что красные, оранжевые и желтые цвета — теплые, а синие и зеленые — холодные, связано только с нашими эмоциями и ассоциациями и не имеет никакого отношения к спектральному составу света от раскаленного тела, зависящему от его температуры, — к тому, что физики называют цветовой температурой.) Если мы будем каким-то способом фильтровать белый свет, удаляя все, кроме узкой спектральной полосы, то получим свет, который называют монохроматическим (см. график на рис. 116 справа).
 Рис. 116. Слева: энергия света (например, солнечного) распределена в широком диапазоне длин волн — примерно от 400 до 700 нанометров. Слабо выраженный пик определяется температурой источника: чем горячее источник, тем больше смещение пика к синему (коротковолновому) концу. Справа: монохроматический свет — это свет, энергия которого сосредоточена в основном в области какой-то одной длины волны. Его можно создать при помощи разнообразных фильтров, лазера или спектроскопа с призмой или дифракционной решеткой.
Рис. 116. Слева: энергия света (например, солнечного) распределена в широком диапазоне длин волн — примерно от 400 до 700 нанометров. Слабо выраженный пик определяется температурой источника: чем горячее источник, тем больше смещение пика к синему (коротковолновому) концу. Справа: монохроматический свет — это свет, энергия которого сосредоточена в основном в области какой-то одной длины волны. Его можно создать при помощи разнообразных фильтров, лазера или спектроскопа с призмой или дифракционной решеткой.
Пигменты
Когда свет падает на некоторый объект, может происходить одно из трех событий: свет может поглощаться, а энергия его превращаться в тепло, как это бывает, когда что-то нагревается на солнце; он может проходить сквозь объект, если, например, на пути солнечных лучей окажется вода или стекло; либо он может отражаться, как в случае зеркала или любого светлого предмета, например куска мела. Часто происходят два или все три события; например, часть света может поглотиться, а часть — отразиться. Для многих объектов относительное количество поглощенного и отраженного света зависит от длины волны. Зеленый лист растения поглощает длинно- и коротковолновый свет и отражает свет промежуточной области спектра, так что при освещении листа солнечными лучами отраженный свет будет иметь выраженный широкий максимум на средних длинах волн (в области зеленого цвета). Красный объект будет иметь свой максимум, тоже широкий, в области длинных волн, как показано на рис. 117. Вещество, которое поглощает часть падающего на него света и отражает остальную часть, называют пигментом. Если какие-то спектральные компоненты в диапазоне видимого света поглощаются лучше, чем другие, пигмент представляется нам окрашенным. Сразу же добавим: какой именно цвет мы видим, зависит не только от длины волн, но также от распределения энергии между разными участками спектра и от свойств нашей зрительной системы. Здесь замешаны как физика, так и биология.
 Рис. 117. Большинство окрашенных объектов отражает свет, энергия которого обычно в некоторых участках видимого спектра больше, чем в других участках. Распределение его энергии, однако, намного шире, чем у монохроматического света. На этом графике показан спектральный состав света, который отразится от красного объекта при его освещении широкополосным (белым) источником.
Рис. 117. Большинство окрашенных объектов отражает свет, энергия которого обычно в некоторых участках видимого спектра больше, чем в других участках. Распределение его энергии, однако, намного шире, чем у монохроматического света. На этом графике показан спектральный состав света, который отразится от красного объекта при его освещении широкополосным (белым) источником.
Зрительные рецепторы
Каждая палочка или колбочка в нашей сетчатке содержит пигмент, поглощающий в каком-то участке спектра лучше, чем в других участках. Поэтому, если бы мы смогли собрать достаточное количество такого пигмента и посмотреть на него, он выглядел бы окрашенным. Зрительный пигмент обладает особым свойством: при поглощении им светового фотона он изменяет свою молекулярную форму и при этом высвобождает энергию, запуская таким образом цепь химических реакций, описанную в главе 3, которые в конце концов приводят к появлению электрического сигнала и к выделению химического медиатора в синапсе. Пигментная молекула в своей новой форме, как правило, обладает совсем иными светопоглощающими свойствами, и если, как это обычно бывает, она поглощает свет хуже, чем в исходной форме, мы говорим, что она «выцветает» под действием света. Затем сложный химический механизм глаза восстанавливает первоначальную конфигурацию пигмента; в противном случае его запас быстро истощился бы. Сетчатка содержит своего рода мозаику из рецепторов четырех типов — палочек и трех типов колбочек (рис. 118). Каждый тип рецепторов содержит свой особый пигмент. Разные пигменты отличаются друг от друга в химическом отношении, а в связи с этим и по способности поглощать свет с различной длиной волн. Палочки ответственны за нашу способность видеть при слабом свете, т.е. за сравнительно грубую разновидность зрения, не позволяющую различать цвета. Палочковый пигмент родопсин обладает наибольшей чувствительностью в области около 510 нм, в зеленой части спектра. Палочки отличаются от колбочек во многих отношениях: они меньше и имеют несколько иное строение, по-иному распределены в разных частях сетчатки и имеют свои особенности в системе связей, образуемых с последующими уровнями зрительного пути. И наконец, по содержащимся в них светочувствительным пигментам три типа колбочек отличаются как друг от друга, так и от палочек.
 Рис. 118. Рецепторы сетчатки образуют мозаику, состоящую из палочек и трех типов колбочек. Данная схема могла бы отображать участок сетчатки в нескольких градусах от центральной ямки, где колбочек больше, чем палочек.
Рис. 118. Рецепторы сетчатки образуют мозаику, состоящую из палочек и трех типов колбочек. Данная схема могла бы отображать участок сетчатки в нескольких градусах от центральной ямки, где колбочек больше, чем палочек.
Пигменты колбочек трех типов имеют пики поглощения в области 430, 530 и 560 нм (рис. 119); поэтому разные колбочки несколько неточно называют соответственно «синими», «зелеными» и «красными». Неточность состоит в том, что 1) эти названия отражают максимумы чувствительности (которые в свою очередь зависят от светопоглощающей способности), а не то, как эти пигменты выглядели бы, если бы на них можно было посмотреть; 2) монохроматический свет с длинами волн 430, 530 и 560 нм будет не синим, зеленым и красным, а фиолетовым, сине-зеленым и желто-зеленым; 3) если бы можно было стимулировать колбочки только одного типа, мы видели бы не синий, зеленый и красный цвета, а, вероятно, фиолетовый, зеленый и желтовато-зеленый. Однако приведенные выше названия колбочек широко распространены, а попытки изменить укоренившуюся терминологию обычно оканчиваются неудачей. Более корректными были бы названия «длинноволновые», «средневолновые» и «коротковолновые», но они затрудняли бы понимание для тех, кто не слишком хорошо знаком со спектром.
 Рис. 119. Спектры поглощения (или кривые спектральной чувствительности) у колбочек трех типов различны. (Ординаты на энергетических кривых и кривых поглощения откладываются в логарифмических единицах, поскольку их значения изменяются в очень широком диапазоне. Поэтому положение оси x произвольно и не соответствует нулевому поглощению).
Рис. 119. Спектры поглощения (или кривые спектральной чувствительности) у колбочек трех типов различны. (Ординаты на энергетических кривых и кривых поглощения откладываются в логарифмических единицах, поскольку их значения изменяются в очень широком диапазоне. Поэтому положение оси x произвольно и не соответствует нулевому поглощению).
Имея максимум поглощения в зеленой области, палочковый пигмент родопсин отражает синие и красные лучи и поэтому выглядит пурпурным. Поскольку в наших сетчатках он присутствует в количествах, достаточных для того, чтобы химики смогли его выделить и можно было на него посмотреть, он издавна получил название зрительного пурпура. Само по себе это нелогично, поскольку «зрительный пурпур» называют так по его видимому цвету, тогда как названия для колбочек («красные», «синие» и «зеленые») соответствуют их относительной чувствительности, т.е. способности поглощать свет. Во избежание путаницы об этом следует помнить. Три типа колбочек имеют широкие зоны чувствительности со значительным перекрыванием, особенно для красных и зеленых колбочек. Свет с длиной волны 600 нм вызовет наибольшую реакцию красных колбочек, пик чувствительности которых расположен при 560 нм; вероятно, он вызовет также некоторую, хотя и более слабую, реакцию колбочек двух других типов. Таким образом, «красная» колбочка реагирует не только на длинноволновый, т.е. красный, свет; она лишь реагирует на него лучше других колбочек. Сказанное относится и к колбочкам других типов. До сих пор я рассматривал физические аспекты цветового зрения: природу света и пигментов, свойства объектов, отражающих свет к нашим глазам, и особенности палочковых и колбочковых пигментов, преобразующих поглощенный свет в электрические сигналы. Интерпретировать эти исходные сигналы как различные цвета — это уже задача мозга. Чтобы лучше дать почувствовать предмет обсуждения, я решил вначале кратко изложить элементарные факты о цветовом зрении, оставив пока в стороне трехсотлетнюю историю установления этих фактов, а также процессы обработки цветовой информации мозгом.
Общие замечания о цвете
Быть может, полезно начать с того, как оперируют с различными длинами волн две сенсорные системы — слуховая и зрительная. Деятельность одной из них приводит к восприятию высоты тона, а другой — к восприятию цвета, но между этими системами есть глубокое различие. Когда я беру на фортепиано аккорд из пяти нот, вы можете выделить отдельные ноты и пропеть каждую из них по отдельности. Ноты не смешиваются в нашем мозгу, но сохраняют свою индивидуальность, в то время как еще со времен Ньютона известно, что при смешивании двух или нескольких световых лучей разного цвета вы не можете выделить компоненты путем простого рассматривания. Небольшое размышление убедит вас в том, что цветовое зрение неизбежно должно быть чувством менее совершенным, чем восприятие тонов. Звук, приходящий в любой данный момент в одно ухо и состоящий из колебаний с разной длиной волны, будет воздействовать на тысячи рецепторов внутреннего уха, каждый из которых настроен на высоту, слегка отличающуюся от настройки соседнего рецептора. Если звук состоит из многих волновых составляющих, информацию будет получать множество рецепторов, все выходные сигналы которых передаются в наш мозг. Богатство слуховой информации определяется способностью мозга анализировать такие комбинации звуков. Совершенно иначе обстоит дело со зрением. Предметом обработки в зрительной системе служит изображение, схватываемое в каждый момент времени набором из миллионов рецепторов. Мы мгновенно воспринимаем сложную сцену. Если при этом мы захотели бы еще обрабатывать длины волн по принципам, используемым во внутреннем ухе, то сетчатка должна была бы иметь не только набор рецепторов, покрывающих всю ее поверхность, но и, скажем, по тысяче рецепторов в каждой отдельной точке, каждый из которых обладал бы максимальной чувствительностью к своей длине волны. Но втиснуть тысячу рецепторов в каждую точку сетчатки физически невозможно; поэтому здесь приходится идти на компромисс. Сетчатка содержит «цветовые» рецепторы трех типов с различной чувствительностью к длине волны в каждой из очень большого числа точек. Таким образом, ценой незначительного ущерба для разрешающей способности бо́льшая часть нашей сетчатки получает некоторую возможность обрабатывать информацию о длинах волн. Мы различаем семь цветов, а не 88 (впрочем, обе цифры следует многократно увеличить с учетом оттенков), но зато каждой из множества тысяч точек видимой сцены будет приписан определенный цвет. Сетчатка не могла бы обладать той способностью к пространственному анализу, которую она имеет, и одновременно обрабатывать информацию о длинах волн столь же изощренно, как слуховая система. Теперь нужно дать читателю представление о том, что означает для нашего цветового зрения обладание тремя типами колбочек. Во-первых, может возникнуть вопрос: если данная колбочка при каких-то длинах волн работает лучше, чем при других, почему бы зрительной системе просто не измерить выход этой колбочки и не вычислить отсюда, каков здесь цвет? Почему бы тогда не иметь колбочки одного типа вместо трех? Да потому, что при одном типе колбочек, скажем красных, вы не смогли бы отличить свет с наиболее эффективной длиной волны в области 560 нм от более яркого света с менее эффективной длиной волны. Необходимо иметь возможность отличать изменения яркости от изменений длины волны. Но предположим, что у вас есть два вида колбочек с перекрывающимися кривыми спектральной чувствительности, например красные и зеленые колбочки. Теперь вы можете определять длину волны простым сравнением выходов колбочек. При коротких волнах сильнее будут реагировать зеленые колбочки; по мере увеличения длины волны реакции тех и других колбочек будут все больше приближаться друг к другу, пока не сравняются; примерно при 580 нм красные начнут отвечать лучше зеленых, и эта разница будет постепенно увеличиваться по мере дальнейшего роста длины волны. Если мы вычтем из кривой чувствительности для одних колбочек кривую для других (это логарифмические кривые, поэтому мы фактически берем отношения величин), то мы получим некоторую кривую, не зависящую от интенсивности света. Таким образом, колбочки двух типов вместе образуют прибор для измерения длины волны. Почему же тогда двух типов рецепторов мало, чтобы полностью объяснить свойства нашего цветового зрения? Двух и в самом деле было бы достаточно, если бы мы имели дело только с монохроматическим светом — если бы мы согласились отказаться от таких вещей, как способность отличать цветной свет от белого. Наше зрение таково, что никакой монохроматический свет с любой длиной волны не выглядит белым. Это было бы невозможно при колбочках только двух типов. В случае красных и зеленых колбочек, продвигаясь от коротких к длинным волнам, мы постепенно переходим от стимуляции только зеленых к стимуляции только красных рецепторов со всеми промежуточными соотношениями между реакциями тех и других. Белый свет, состоящий, по существу, из смеси всех волн, должен в определенной мере стимулировать и красные, и зеленые колбочки. Таким образом, если монохроматический свет будет иметь длину волны, дающую то же соотношение реакций, то он будет неотличим от белого. Именно так обстоит дело при наиболее распространенной форме цветовой слепоты, когда человек имеет только два вида колбочек: независимо от того, какой из трех пигментов отсутствует, всегда найдется свет с какой-то длиной волны, неотличимый от белого. (Эти люди обладают дефектами цветового восприятия, но, конечно, не являются полностью цветнослепыми.) Чтобы иметь цветовое зрение, подобное нашему, необходимо и достаточно иметь колбочки трех типов. Вывод о том, что у нас действительно именно три типа колбочек, был впервые сделан при исследовании особенностей цветового зрения человека, в результате ряда дедуктивных умозаключений, делающих честь человеческому интеллекту. Теперь мы можем лучше понять, почему палочки не участвуют в восприятии цвета. При промежуточных уровнях освещенности могут функционировать как палочки, так и колбочки, но нервная система (если не считать редких искусственных ситуаций), по-видимому, не занимается вычитанием палочковых влияний из колбочковых. Колбочки сравнивают друг с другом, а палочки работают сами по себе. Если вы хотите убедиться в том, что палочки не передают информацию о цвете, проснитесь лунной ночью и оглядитесь. Хотя форму предметов вы сможете видеть довольно хорошо, цвета будут полностью отсутствовать. Удивительно, как мало людей осознают, что при слабом свете они обходятся без цветового зрения. Увидим ли мы данный объект белым или цветным, определяется главным образом (но не всецело) тем, какие из трех типов колбочек активируются. Цвет — это результат неодинаковой стимуляции колбочек разного типа. Понятно, что свет с широкой спектральной кривой, например от солнца или от свечи, будет стимулировать колбочки всех трех типов (возможно, почти одинаково), и тогда ощущение окажется лишенным цвета, или «белым». Если бы нам удалось стимулировать колбочки только одного вида (что с помощью света сделать нелегко из-за перекрывания кривых поглощения), то результатом, как уже говорилось, был бы яркий цвет — фиолетовый, зеленый или красный в зависимости от вида стимулируемых колбочек. То, что максимум чувствительности тех колбочек, которые мы называем «красными», соответствует длине волны света, видимого нами как зеленовато-желтый (560 нм), связано, по-видимому, с тем, что такой свет возбуждает как зеленые, так и красные колбочки — из-за перекрывания кривых их спектральной чувствительности. Используя свет с большей длиной волны, мы можем более эффективно стимулировать красные колбочки по сравнению с зелеными. Графики на рис. 120 резюмируют цветовые ощущения, возникающие при активации разных сочетаний колбочек светом различного спектрального состава. Первый и два последних примера должны убедительно показать, что ощущение «белого» цвета — результат примерно одинаковой стимуляции колбочек всех трех типов — может быть вызвано многими различными способами: и воздействием широкополосного света, и с помощью смеси узких спектральных полос, например желтого света с синим или красного с сине-зеленым. Два световых луча называют дополнительными, если их волновой состав и интенсивность подобраны так, что при смешении они дают ощущение «белого». В двух последних примерах синий и желтый, так же как красный с длиной волны 640 нм и сине-зеленый цвета являются дополнительными.
 Рис. 120. Верхний график — «чувствительность колбочек» — повторяет график, приведенный на рис. 119. Далее указано, какие колбочки будут активироваться различными смесями окрашенного света и какие будут возникать ощущения.
Рис. 120. Верхний график — «чувствительность колбочек» — повторяет график, приведенный на рис. 119. Далее указано, какие колбочки будут активироваться различными смесями окрашенного света и какие будут возникать ощущения.
Теории цветового зрения
Все сказанное выше о зависимости видимого цвета от стимуляции тех или иных колбочек основано на исследованиях, начатых Ньютоном в 1704 году и продолжающихся до сих пор. Изобретательность, которую проявил Ньютон в своих экспериментах, трудно переоценить: в работе, посвященной цвету, он при помощи призмы расщеплял белый свет; воссоединял его компоненты второй призмой, вновь получая белый свет; изготовил волчок с цветовыми секторами, при вращении которого опять-таки получался белый цвет. Эти открытия привели к осознанию того, что обычный свет состоит из непрерывного ряда лучей с различными длинами волн. В XVIII столетии постепенно выяснилось, что всякий цвет можно получить путем смешения трех цветных компонентов в надлежащих пропорциях при условии, что длины их волн достаточно отличаются друг от друга. Представление о том, что любой цвет может быть «составлен» путем манипулирования тремя управляющими факторами (в данном случае путем изменения интенсивности трех различных лучей) получило название трихроматичности. В 1802 году Томас Юнг выдвинул четкую и простую теорию, объясняющую трихроматичность: он предположил, что в каждой точке сетчатки должны существовать по меньшей мере три «частицы» — крошечные структуры, чувствительные соответственно к красному, зеленому и фиолетовому. Длительный временной интервал между Ньютоном и Юнгом трудно объясним, но различные «дорожные препятствия» вроде, например, того факта, что желтая и синяя краски, смешиваясь, дают зеленую, не способствовали, конечно, ясности мышления. Решающие эксперименты, прямо и недвусмысленно подтвердившие, наконец, идею Юнга о том, что цвет должен определяться мозаикой трех видов детекторов в сетчатке, были проведены в 1959 году: Джордж Уолд и Пол Браун в Гарварде и Эдвард Мак-Никол и Уильям Маркс в Университете Джонса Гопкинса изучали под микроскопом способность отдельных колбочек поглощать свет с различной длиной волны и обнаружили три и только три типа колбочек. До этого ученые прилагали все усилия, используя менее прямые методы, и за несколько столетий фактически пришли к такому же результату, доказав теорию Юнга о необходимости именно трех типов колбочек и оценив их спектральную чувствительность. Применялись в основном психофизические методы: ученые выясняли, какие цветовые ощущения вызывают различные смеси монохроматических лучей, как влияет на цветовое зрение избирательное обесцвечивание рецепторов под действием монохроматического света, а также исследовали цветовую слепоту. Изучение эффектов смешения цветов необычайно интересно — настолько его результаты удивительны и противоречат интуиции. Никто без предварительного знания не угадал бы разнообразные явления, иллюстрируемые на рис. 120 и 121, — например, не мог бы предсказать, что два пятна, ярко-синее и ярко-желтое, при наложении друг на друга сольются в белый цвет, неотличимый на глаз от цвета мела, или что зеленый и красный спектральные цвета при их объединении дадут желтый, почти неотличимый от монохроматического желтого цвета.
 Рис. 121. С помощью трех диапроекторов и трех фильтров на экран проецируются три перекрывающихся пятна (красное, зеленое и синее). Красное и зеленое при наложении дают желтый цвет, синее и зеленое — бирюзовый, красное и синее — пурпурный, а все три вместе — белый цвет.
Рис. 121. С помощью трех диапроекторов и трех фильтров на экран проецируются три перекрывающихся пятна (красное, зеленое и синее). Красное и зеленое при наложении дают желтый цвет, синее и зеленое — бирюзовый, красное и синее — пурпурный, а все три вместе — белый цвет.
Прежде чем обсуждать другие теории цвета, нужно сообщить ряд дополнительных сведений о разнообразии цветов, которое эти теории призваны объяснить. Какие существуют цвета помимо цветов радуги? По моему мнению, имеются три вида таких цветов. Один вид — пурпурные, которые отсутствуют в радуге, но появляются при одновременной стимуляции красных и синих колбочек, т.е. при смешении длинно- и коротковолнового, или, грубо говоря, красного и синего света. Если к смеси спектрального красного и спектрального синего света — к пурпурному — мы добавим надлежащее количество зеленого, то мы получим белый цвет; поэтому мы говорим, что зеленый и пурпурный являются дополнительными. Можно, если угодно, представить себе круговую шкалу, включающую все цвета спектра от красного через желтый и зеленый до синего и фиолетового с последующим переходом к пурпурным цветам — сначала к синевато-пурпурному, затем к красновато-пурпурному и наконец опять к красному. Можно расположить эти оттенки так, чтобы дополнительные цвета располагались друг против друга. Понятие основных цветов не вписывается в эту схему: если определить основные цвета в соответствии с тремя типами рецепторов, то мы выделим зеленовато-желтый, зеленый и фиолетовый, т.е. оттенки, вряд ли согласующиеся с представлением о трех чистых базовых цветах. Но если под основными подразумевать три цвета, из которых можно получить любой другой оттенок, то упомянутые три цвета этому критерию удовлетворяют, как, впрочем, и любые другие три достаточно далеко отстоящие друг от друга цвета. Таким образом, ничто из сказанного выше не обосновывает представление о трех единственных основных цветах. Второй тип цвета получается от добавления белого к любому цвету спектра или к пурпурному; мы говорим, что такое добавление «разбавляет» цвет, делает его бледнее — на профессиональном языке говорят, что белый уменьшает насыщенность цвета. Для подбора двух идентичных цветов мы должны сделать их одинаковыми по тону и насыщенности (выбрав, например, соответствующее положение на цветовом круге и затем добавив нужное количество белого), а потом уравнять по интенсивности. Таким образом, мы можем определить некоторый цвет, задав длину волны света (или в случае пурпурного — дополнительного к нему цвета), относительное содержание белого света и число, характеризующее интенсивность. Математически эквивалентная возможность определения цвета состоит в задании трех чисел, представляющих относительные влияния света на три типа колбочек. В любом случае необходимы три числа. Типичным примером цвета третьего типа, не укладывающегося в приведенные выше объяснения, является коричневый. Я вернусь к нему позднее. Герман Гельмгольц принял и отстаивал теорию Юнга, которая приобрела известность как теория Юнга—Гельмгольца. Между прочим, именно Гельмгольц объяснил, наконец, феномен, упомянутый в начале этой главы и заключающийся в том, что смесь желтой и синей красок дает зеленую. Вы можете легко убедиться, насколько это отличается от смешения желтого и синего света, проделав следующий опыт, для которого вам понадобятся всего лишь два диапроектора и немного желтого и синего целлофана. Сначала прикрепите желтый целлофан к линзе одного проектора, а синий — к линзе другого и наложите проецируемые изображения друг на друга. Отрегулировав относительные интенсивности, вы получите в зоне перекрывания чистый белый свет. Этот вид смешения цветов мы уже рассматривали; как мы тогда объяснили, белый свет возникает из-за того, что совместное воздействие желтого и синего света активирует все три системы колбочек с той же относительной эффективностью, что и широкополосный, или белый, свет. Теперь выключите один проектор и расположите оба фильтра перед другим; вы получите зеленый цвет. Чтобы понять, отчего так происходит, мы должны знать, что синий целлофан поглощает из белого света длинноволновую часть, т.е. желтый и красный, а остальную, которая выглядит синей, пропускает, в то время как желтый фильтр поглощает в основном синюю часть, а остальную, кажущуюся желтой, пропускает. Схема на рис. 122 показывает спектральный состав света, пропускаемого каждым фильтром. Обратите внимание на то, что в обоих случаях пропускаемый свет далек от монохроматического. Желтый свет — это не узкополосный спектральный желтый, а смесь спектрального желтого с более короткими зелеными, более длинными оранжевыми и красными волнами. Аналогичным образом, синий — это спектральный синий с примесью зеленого и фиолетового. Почему же в таком случае мы видим только желтый или только синий цвет? Дело в том, что ощущение желтого — результат одинаковой стимуляции красных и зеленых колбочек без какого-либо воздействия на синие колбочки; такую стимуляцию можно осуществлять как спектральным желтым (монохроматическим светом с длиной волны 580 нм), так и более широким волновым «мазком», который обычно свойствен пигментам, — нужно лишь, чтобы ширина спектра не была чрезмерно велика и спектр не содержал коротких волн, стимулирующих синие колбочки. Аналогичным образом, спектральный синий свет оказывает приблизительно такое же воздействие, как синий плюс зеленый плюс фиолетовый. Теперь при использовании двух фильтров, расположенных один перед другим, мы получим то, что́ пропускают оба фильтра, т.е. зеленые лучи. Именно в этой области перекрываются приведенные на рис. 128 графики для широкополосного синего и желтого света. То же самое происходит с красками: желтая и синяя краски вместе поглощают весь свет, кроме зеленых участков, которые отражаются. Отметим, что если бы мы использовали в нашем опыте монохроматические желтый и синий фильтры, расположив их один перед другим, они не пропустили бы ничего. Смешивание происходит только потому, что свет, пропускаемый или отражаемый красящими веществами, имеет широкополосный спектральный состав.
 Рис. 122. Синий фильтр пропускает довольно широкую спектральную полосу с центром в области 480 нм, а желтый — такую же полосу с центром в области 580 нм. Оба фильтра вместе пропускают только общие для них лучи — свет в достаточно узкой полосе с центром около 530 нм, что дает зеленый цвет.
Рис. 122. Синий фильтр пропускает довольно широкую спектральную полосу с центром в области 480 нм, а желтый — такую же полосу с центром в области 580 нм. Оба фильтра вместе пропускают только общие для них лучи — свет в достаточно узкой полосе с центром около 530 нм, что дает зеленый цвет.
Резюмируем это многословное объяснение того, почему «желтый плюс синий равен зеленому», следующим кратким утверждением о цвете и красителях: два фильтра, расположенные один перед другим, или две смешанные краски совместно поглощают из белого света все, кроме волн средней длины, т.е. зеленого цвета. Почему я обсуждаю здесь этот феномен? Отчасти потому, что это объясняет драматичный и сенсационный результат смешения желтого и синего с получением зеленого, но еще больше из-за исторической важности этого результата для подтверждения нашего понимания цветового зрения. Феномен этот физический; он связан с цветовым зрением и биологией примерно так же, как с ними связаны перекрещивание поляроидов и получение черного или добавление к кислоте синего лакмуса и получение красного, — говоря коротко, никак. И все-таки мысль о связи смешения цветов с цветовым зрением еще продолжает смущать многих, и это обусловлено представлением о том, что красный, желтый и синий — основные цвета, а зеленый — нет. Если какой-либо набор цветов и можно признать основным, так это четыре цвета — красный, синий, желтый и зеленый. Как мы убедимся в разделе, посвященном теории Геринга, основание, с которым все четыре цвета могут претендовать на звание основных, мало связано с тремя типами колбочек и значительно больше — с последующей переработкойинформации в сетчатке и в мозгу. (Этим нисколько не обесценивается знание живописцем того факта, что всего тремя красками можно имитировать большинство цветовых оттенков. Но даже мастер в своей области может ошибиться. В одной книге, посвященной ткацкому делу, в главе, излагающей теорию цвета, я нашел утверждение, что если вы смешаете в ткани желтые и синие нити, то получите зеленый цвет. На самом же деле получится серый цвет — по биологическим причинам.)
Цветовая слепота
Из работ Дж. Уолда, У. Раштона и многих других мы знаем, что в основе обычных форм цветовой слепоты, имеющейся примерно у 8 процентов мужчин, лежит отсутствие или нехватка одного или нескольких типов колбочек. Число возможных комбинаций отсутствия или количественного недостатка тех или иных колбочек делает цветовую слепоту весьма сложным объектом исследования. Иногда цветовая слепота возникает в левом или правом поле зрения после локального инсульта в контралатеральном или ипсилатеральном полушарии. При этом, вероятно, повреждается какая-то высшая корковая зрительная зона, расположенная выше стриарной коры и зоны 18, — зона, названная V4 Семиром Зеки из Университетского колледжа. Об этих высших зонах мы поговорим в главе 10.
Теория Геринга
Параллельно теории цвета Юнга—Гельмгольца возникла и до недавнего времени казалась с ней несовместимой вторая научная школа. Эвальд Геринг (1834–1918) интерпретировал результаты смешения цветов, предположив, что в глазу и/или мозгу существуют три оппонентных процесса: один для ощущения красного и зеленого, другой для желтого и синего и третий, качественно отличный от двух первых, — для черного и белого. Геринга поразило отсутствие (их невозможно даже представить себе!) цветов, которые можно было бы описать как желтовато-синий или красновато-зеленый, а также «взаимное уничтожение» синего и желтого или красного и зеленого при их смешении в надлежащих пропорциях — цвет при этом полностью исчезает, т.е. возникает ощущение белого цвета. Геринг рассматривал красно-зеленый и желто-синий процессы как независимые в том смысле, что смесь синего и красного дает синевато-красный, или пурпурный; аналогично, смесь красного и желтого дает оранжевый, смесь зеленого и синего — синевато-зеленый, а смесь зеленого и желтого — зеленовато-желтый. В системе Геринга желтый, синий, красный и зеленый могут считаться «основными» цветами. Смотря на оранжевое, каждый может представить его себе как результат смешения красного и желтого, но никому не удается, глядя на красное или синее, видеть в нем результат смешения каких-либо других цветов. (Возникающее у некоторых людей ощущение, будто зеленый выглядит как желтый с добавкой синего, обусловлено, вероятно, их детским жизненным опытом, связанным с наборами красок.) Многим казалось, что представления Геринга о сине-зеленом и желто-синем процессах опираются всего лишь на интуитивные впечатления о цвете. Но поражает, насколько хорошо согласуются мнения людей, которых просили указать точку спектра, где представлен чистый синий цвет без какой-либо кажущейся примеси зеленого или желтого. То же самое можно сказать о желтом и зеленом цветах. Что касается красного цвета, то оценка испытуемых вновь совпадает, но в этом случае они настаивают на том, что нужно добавить немного фиолетового, чтобы устранить чуть заметную желтизну длинноволнового света. [Именно такой субъективный красный при добавлении к зеленому дает белый; обычный (спектральный) красный, добавленный к зеленому, дает желтый.] Мы можем сравнить желто-синий и красно-зеленый процессы Геринга с двумя приборами вроде старинных вольтметров, стрелка одного из которых отклоняется влево при регистрации желтого и вправо при регистрации синего, а другой прибор ведет себя точно так же по отношению к паре «красный-зеленый». Цвет объекта при этом можно описать показаниями двух приборов. Третий антагонистический процесс Геринга (его можно представить себе как третий вольтметр) регистрирует соотношение черного и белого. Геринг понимал, что ощущение черного и серого порождается не просто отсутствием света, поступающего от некоторого объекта или поверхности, а возникает тогда и только тогда, когда от объекта приходит меньше света, чем в среднем от окружающих областей. Ощущение белого возникает только в том случае, если фон темнее и отсутствует цвет (это я уже обсуждал в главе 3 на примере выключенного телевизора). По теории Геринга черно-белый процесс предполагает пространственное сравнение или вычитание отражающих способностей, в то время как его желто-синий и красно-зеленый процессы происходят в одном определенном участке поля зрения и не связаны с окружением. (Геринг, несомненно, знал о взаимодействии соседних цветов, но его теория цвета, как она сформулирована в его поздних работах, не включает эти явления.) Мы уже видели, что черное и белое действительно представлены в сетчатке и в мозгу пространственно разделенными процессами возбуждения и торможения (on—off), которые в буквальном смысле слова антагонистичны. Теория Геринга позволила объяснить не только все спектральные цвета и уровни насыщенности, но и такие цвета, как коричневый и оливково-зеленый, которые отсутствуют в радуге и даже не могут быть воспроизведены ни в одной из классических психофизических процедур смешения цветов, в которых мы с помощью диапроектора отбрасываем световые пятна на темный экран. Мы получим коричневый цвет лишь в том случае, если желтое или оранжевое световое пятно будет окружено в среднем более ярким светом. Возьмите любую коричневую поверхность, посмотрите на нее через свернутый трубкой кусок черной бумаги, чтобы исключить все ее окружение, и вы увидите желтый или оранжевый цвет. Мы можем считать коричневый цвет смесью черного, получаемого только в условиях пространственного контраста, с оранжевым или желтым. По терминологии Геринга, при этом работают по меньшей мере две системы — черно-белая и желто-синяя. Теория Геринга о трех оппонентных системах — красно-зеленой, желто-синей и черно-белой — в его время и еще полстолетия рассматривали как альтернативную по отношению к трехкомпонентной («красный, зеленый, синий») теории Юнга — Гельмгольца. Сторонники каждой из них были, как правило, весьма фанатичны и зачастую чрезмерно эмоциональны. Физики обычно примыкали к лагерю Юнга — Гельмгольца, быть может потому, что их привлекали количественные аргументы (такие, например, как системы линейных уравнений) и отталкивали доводы, связанные с чистотой цветов. Психологи часто были на стороне Геринга, вероятно в связи с тем, что им приходилось иметь дело с более широким разнообразием психофизических феноменов. Теория Геринга, казалось, содержала доводы в пользу либо четырех типов рецепторов (красный, зеленый, желтый и синий), либо трех (черно-белый, желто-синий и красно-зеленый); оба варианта противоречили накапливавшимся данным, которые подкрепляли исходную гипотезу Юнга. Ретроспективно можно сказать, как отмечают современные психофизики Лео Гурвич и Доротея Джеймсон, что одна из трудностей была связана с отсутствием до 1950-х годов каких-либо прямых физиологических данных о тормозных механизмах в сенсорных системах. Такие данные появились лишь тогда, когда стала возможной регистрация активности одиночных нейронов. Представив себе вольтметры, измеряющие положительные величины отклонением вправо, а отрицательные — влево, вы сможете понять, почему теория Геринга предполагает наличие тормозных механизмов. Желтый и синий цвета взаимно антагонистичны; смешиваясь, они уничтожают друг друга, а если стрелка красно-зеленой системы тоже указывает на нуль, то цвет отсутствует. Геринг в известном смысле на пятьдесят лет опередил свое время. Как это случалось и раньше в истории науки, две теории, на протяжении десятилетий казавшиеся несовместимыми, обе оказались верны. В конце прошлого столетия никто не мог предположить, что представления Юнга — Гельмгольца окажутся верными для рецепторного уровня, а идеи Геринга об оппонентных процессах — для последующих уровней зрительной системы. Теперь стало ясно, что эти две формулировки не исключают друг друга: обе они предполагают наличие системы с тремя переменными — это три типа колбочек в теории Юнга—Гельмгольца и три измерительных прибора или процесса в теории Геринга. Что нас сегодня изумляет, так это то, что Геринг, опираясь на столь ограниченный фактический материал, сумел сформулировать теорию, так хорошо согласующуюся с нейронной организацией центральных механизмов цветового зрения. Тем не менее специалисты по цветовому зрению все еще разделены на два лагеря: одни считают Геринга пророком, а другие видят в упомянутом соответствии лишь случайную удачу. Я наверняка наживу врагов среди тех и других, так как занимаю нейтральную позицию и только чуть-чуть склоняюсь в пользу первого мнения.
Цвет и пространство
В главе 3 мы видели, что восприятие объекта как белого, черного или серого зависит от его относительной способности отражать свет по сравнению с другими предметами, находящимися в поле зрения. Таким образом, свойства широкополосных клеток нижних уровней зрительной системы — ганглиозных клеток сетчатки и клеток коленчатого тела — позволяют в значительной мере объяснить восприятие черного, белого и серого: именно такое сравнение они осуществляют при помощи своих рецептивных полей с центром и периферией. Несомненно, именно в этом состоит третий, пространственно оппонентный черно-белый процесс Геринга. То, что пространственная переменная важна и для восприятия других цветов, начали впервые осознавать еще столетие назад; однако аналитический подход к этому вопросу стал разрабатываться только в последние десятилетия, в основном усилиями таких психофизиков, как Лео Гурвич и Доротея Джеймсон, Дин Джадд и Эдвин Лэнд. Лэнд с его глубоким интересом к проблемам освещения и фотографии был, естественно, заинтригован неспособностью фотокамеры компенсировать различия в источниках света. Если фотопленка сбалансирована так, что изображение белой рубашки выглядит белым при свете вольфрамовых ламп накаливания, то та же рубашка под синим небом будет светло-голубой; если же пленка предназначена для естественного света, рубашка при обычном электрическом свете будет розовой. При изготовлении хорошей цветной фотографии мы должны учитывать не только интенсивность света, но и его спектральный состав — будет ли свет голубоватым или красноватым. Если мы знаем это, мы сможем установить выдержку и диафрагму с учетом интенсивности и подобрать пленку или фильтры с учетом цветового баланса. В отличие от фотокамеры наша зрительная система делает все это автоматически; она настолько хорошо решает эту задачу, что обычно мы даже не осознаем, что такая проблема существует. Белая рубашка выглядит белой, несмотря на большие сдвиги в спектральном составе света при переходе от освещения солнцем в зените к освещению заходящим солнцем, вольфрамовой или флуоресцентной лампой. То же постоянство сохраняется для окрашенных объектов, и этот феномен применительно к цветному и белому называют константностью цвета. Хотя константность известна уже давно, демонстрации Лэнда в 50-х годах явились большим сюрпризом даже для нейрофизиологов, физиков и большинства психологов.
 Рис. 123. Во многих своих экспериментах Эдвин Лэнд использовал мозаику из цветных бумажных полосок в стиле Мондриана. Целью экспериментов было доказательство того, что воспринимаемые цвета сохраняют поразительное постоянство, несмотря на заметные изменения в соотношении красных, зеленых и синих лучей, использованных для освещения мозаики.
Рис. 123. Во многих своих экспериментах Эдвин Лэнд использовал мозаику из цветных бумажных полосок в стиле Мондриана. Целью экспериментов было доказательство того, что воспринимаемые цвета сохраняют поразительное постоянство, несмотря на заметные изменения в соотношении красных, зеленых и синих лучей, использованных для освещения мозаики.
Каковы же эти демонстрации? В типичном эксперименте мозаику из прямоугольных кусков бумаги разного цвета, напоминающую рисунки Мондриана, освещают тремя диапроекторами, один из которых снабжен красным, другой зеленым и третий синим фильтрами. Каждый проектор имеет регулируемый источник света, так что интенсивность его можно изменять в широких пределах. В остальном комната должна быть полностью затемнена. Если все три проектора установить на среднюю интенсивность, то цвета будут выглядеть примерно так же, как при дневном свете. Удивительно, что точная установка как будто не имеет значения. Выберем зеленый участок мозаики и фотометром, точно измерим интенсивность приходящего от него света, когда включен только один проектор. Затем повторим измерение со вторым проектором, а потом с третьим. Это даст нам три числа, характеризующих свет, отражаемый при включении всех трех проекторов. Выберем теперь другой участок, например оранжевый, и отрегулируем поочередно интенсивность каждого проектора так, чтобы показания фотометра для оранжевого участка совпадали с теми, которые мы получили ранее для зеленого. Таким образом, при включении трех проекторов свет, приходящий теперь от оранжевого участка, идентичен по составу тому, который минутой ранее приходил от зеленого участка. Что мы ожидаем увидеть? Рассуждая примитивным образом, мы скажем, что оранжевый участок превратился в зеленый. Но он все-таки выглядит оранжевым — его цвет даже нисколько не изменился. Мы можем повторить этот эксперимент с любыми двумя участками. Вывод таков: не столь важно, на какую интенсивность света установлены три проектора, пока от каждого из них поступает какой-то свет. Почти в любом случае мы заметим лишь очень малые изменения воспринимаемого цвета. Такие эксперименты убедительно показали, что ощущение, возникающее в какой-либо части поля зрения, зависит как от света, приходящего от этой части, так и от света, приходящего от остальных участков. Иначе как мог бы свет одинакового спектрального состава вызывать в одном случае ощущение зеленого, а в другом — ощущение оранжевого? Принцип, применимый к черному, белому и серому и столь четко сформулированный Герингом, оказывается верным и в отношении цвета. Для цвета мы имеем оппонентность не только локальную (красный/зеленый и желтый/синий), но также и пространственную: красный/зеленый в центре против красного/зеленого на периферии и аналогичную оппонентность для желтого/синего. В 1985 году Дэвиду Инглу в лаборатории Лэнда удалось в аквариуме с подводной мозаикой из разноцветных прямоугольников обучить золотую рыбку подплывать к участку определенного цвета. Он обнаружил, что рыбка плывет к одному и тому же цвету, например синему, независимо от спектрального состава света: она, как и мы, выбирает синий участок, даже если свет от него идентичен по составу свету, который в предыдущей пробе при ином источнике света приходил от желтого лоскута, отвергаемого рыбкой. Таким образом, рыба тоже выбирает участок поверхности по его собственному цвету, а не по спектральному составу отражаемого им света. Это означает, что феномен константности цвета не следует считать каким-то усовершенствованием, недавно добавленным в ходе эволюции к цветоощущению некоторых высших млекопитающих, в том числе и человека; наличие его у рыб указывает на то, что это примитивный, весьма общий аспект цветового зрения. Очень заманчиво (и довольно несложно) было бы проверить, обладают ли той же способностью насекомые с цветовым зрением. Я думаю, что это именно так. Лэнд и его группа (среди прочих Дж. Мак-Кэнн, Н. Доу, М. Бёрнс и X. Перри) разработали несколько процедур для предсказания видимого цвета некоторого объекта по спектрально-энергетическому составу света от всех точек поля зрения, но без какой-либо информации о источнике света. Расчет заключается в том, что для каждого из трех отдельных проекторов определяют отношение света, приходящего от места, цвет которого нужно предсказать, к усредненному свету, приходящему от окружения. (Площадь «окружения», которую нужно учитывать, в разных вариантах теории Лэнда различна. В новейшем варианте предполагается, что влияние окружающих участков уменьшается с расстоянием.) Получаемая тройка чисел — отношения берутся для каждого проектора — однозначно определяет цвет данного места. Любому цвету, таким образом, можно поставить в соответствие некоторую точку в трехмерном пространстве, координатными осями которого будут три отношения, полученные для красного, зеленого и синего света. Чтобы сделать формулировку как можно более реалистичной, три источника света подбираются в соответствии с кривыми спектральной чувствительности трех типов колбочек человека.
 Рис. 124. В экспериментах Дэвида Ингла золотая рыбка обучалась подплывать за вознаграждение — кусочек печенки — к полоске заданного цвета. Она плывет, например, к желтой полоске независимо от соотношения интенсивностей света от трех проекторов. Такое поведение поразительно сходно с константностью восприятия цвета у человека.
Рис. 124. В экспериментах Дэвида Ингла золотая рыбка обучалась подплывать за вознаграждение — кусочек печенки — к полоске заданного цвета. Она плывет, например, к желтой полоске независимо от соотношения интенсивностей света от трех проекторов. Такое поведение поразительно сходно с константностью восприятия цвета у человека.
То, что таким образом можно вычислить цвет, означает наличие константности цвета, поскольку для каждого проектора подсчитывается соотношение света от одной области к свету от усредненного окружения. Точная установка силы света в проекторах более не важна: единственным условием остается то, что мы должны иметь какой-то свет от каждого проектора — иначе нельзя будет вычислить никакого отношения. Одно из следствий всего этого заключается в том, что для появления цвета необходимы различия в спектральном составе света в пределах поля зрения. Для восприятия цвета нам нужны цветовые границы, точно так же как нужны границы освещенности для восприятия черного и белого. Вы можете легко убедиться в этом, снова использовав два диапроектора. Поставьте перед одним из проекторов красный фильтр (подойдет красный целлофан) и осветите любую группу предметов. Я предпочитаю брать белую или желтую рубашку и яркий красный галстук. При таком освещении ни рубашка, ни галстук не выглядят вполне красными: и то и другое кажется розоватым и как бы полинявшим. Теперь осветите ту же комбинацию предметов вторым проектором, прикрыв его синим целлофаном. Рубашка будет выглядеть бледно-голубоватой, а галстук — черным: красные объекты не отражают коротких световых волн. Вернитесь к красному проектору и убедитесь еще раз, что галстук не выглядит особенно красным. Теперь добавьте синий проектор. Вы знаете, что при добавлении синего света вы не получите от галстука какого-либо дополнительного отражения — вы только что это продемонстрировали, — но при включении синего проектора красный галстук внезапно засверкает хорошим ярким красным цветом. Это убедит вас в том, что галстук делают красным не только лучи, приходящие от него самого. Эксперименты со стабилизированными цветовыми границами согласуются с представлением о том, что вообще для видения цвета необходимы различия на границах. Альфред Ярбус, имя которого упоминалось в главе 4 в связи с движением глаз, в 1962 году показал, что если смотреть на синее пятно, окруженное красным фоном, стабилизация границы пятна на сетчатке приводит к его исчезновению: синее пропадает, и все, что можно видеть, — это красный фон. Стабилизация границ на сетчатке явно делает их неэффективными, а без них исчезает и цвет. Эти психофизические доказательства того, что для восприятия цвета нужны различия в спектральном составе света от разных участков поля зрения, указывают на возможное наличие в наших сетчатках или мозгу клеток, чувствительных к цветовым границам. Этот довод аналогичен тому, который приводился нами в главе 4 по поводу восприятия черных или белых объектов. Если на каком-то уровне нашей зрительной системы информация о цвете передается только на линиях цветового контраста, то клетки с рецептивными полями, лежащими целиком внутри областей однородного цвета, будут бездействовать. Результат — экономия при обработке информации. Таким образом, от передачи сведений о цвете лишь на границах мы получаем два преимущества: во-первых, цвет не меняется при изменениях освещения, так что мы узнаём о свойствах рассматриваемых объектов без искажений, вносимых источником света; во-вторых, информация обрабатывается экономным способом. Теперь мы можем задаться вопросом: почему система эволюционировала именно таким образом? Была ли главным фактором эволюции необходимость константности цвета, а экономия — лишь сопутствующей выгодой? Или, наоборот, экономия играла ведущую роль, а константность — второстепенную. Второе предположение многим может показаться более убедительным: вряд ли эволюция могла предвидеть появление ламп накаливания или флуоресцентного освещения, да и наши рубашки вовсе не были такими белыми, пока не появились современные моющие средства.
Физиология цветового зрения: ранние результаты
Первая физиологическая информация на клеточном уровне была получена спустя 250 лет после Ньютона в исследованиях шведско-финско-венесуэльского физиолога Гуннара Светихина, который в 1956 году на костистой рыбе осуществил внутриклеточную регистрацию активности нейронов сетчатки — сначала он принял их за колбочки, но они оказались горизонтальными клетками. На освещение сетчатки эти клетки отвечали только медленными потенциалами (потенциалов действия не наблюдалось). Как показано на рис. 125, Светихин обнаружил три типа клеток: первый тип, названный им L-клетками, гиперполяризовался при световой стимуляции независимо от спектрального состава света; второй тип, названной r-g-клетками, гиперполяризовался волнами малой длины с максимумом ответа на зеленый свет и деполяризовался волнами большой длины с максимумом ответа на красный свет; третий тип, названный с учетом теории Геринга y-клетками, отвечал по типу клеток r-g, но с максимумом гиперполяризации на синий и максимумом деполяризации на желтый свет. У клеток r-g и y-b белый свет вызывал лишь слабые и быстро затухающие ответы, как и следовало ожидать ввиду широкополосного спектрально-энергетического состава белого света. Кроме того, у клеток обоих этих типов, которые мы можем назвать оппонентно-цветовыми клетками, свет с некоторой промежуточной длиной волны, называющейся точкой перекреста, не вызывал никакой реакции. Поскольку эти клетки реагируют на окрашенный, но не на белый свет, они, вероятно, связаны с цветовыми ощущениями.
 Рис. 125. Гуннар Светихин и Эдуард Мак-Никол регистрировали реакции горизонтальных клеток на цвет у костистой рыбы. Отклонения вниз от серой линии соответствуют гиперполяризации, а отклонения вверх — деполяризации.
Рис. 125. Гуннар Светихин и Эдуард Мак-Никол регистрировали реакции горизонтальных клеток на цвет у костистой рыбы. Отклонения вниз от серой линии соответствуют гиперполяризации, а отклонения вверх — деполяризации.
В 1958 году Рассел де Валуа и его сотрудники зарегистрировали ответы, поразительно сходные со светихинскими, от клеток наружного коленчатого тела макака. Ранее с помощью поведенческих тестов де Валуа показал, что цветовое зрение макака и человека почти одинаково; например, соотношение, в котором нужно смешать два цветных луча, чтобы получить третий цвет, почти идентично у обоих видов. Поэтому можно думать, что макаки и люди обладают сходными механизмами на нижних уровнях зрительной системы, и мы, по-видимому, вправе сравнивать психофизику цвета у человека с физиологией у макака. Де Валуа обнаружил, что многие клетки коленчатого тела активировались рассеянным монохроматическим светом в диапазоне от одного конца спектра до точки перекреста, где реакция отсутствовала, и подавлялись светом во втором диапазоне — от точки перекреста до другого конца спектра. И вновь аналогия с цветовыми процессами Геринга была полной: де Валуа выявил оппонентно-цветовые клетки двух типов, красно-зеленые и желто-синие; у каждого типа смешение двух световых волн, длины которых на волновой шкале располагались симметрично относительно точки перекреста, приводило к взаимному погашению реакций точно так же, как в восприятии добавление синего к желтому или зеленого к красному порождает белый. Результаты де Валуа особенно хорошо согласовались с формулировками Геринга, поскольку две группы цветовых клеток имели максимумы реакции и точки перекреста точно в тех местах спектральной шкалы, чтобы одна группа могла отражать «желто-синие» свойства падающего света, а другая группа — «красно-зеленые».
 Рис. 126. В типичном рецептивном поле типа 1 центр посылает возбуждающие сигналы от красных колбочек, а периферия — тормозящие сигналы от зеленых колбочек.
Рис. 126. В типичном рецептивном поле типа 1 центр посылает возбуждающие сигналы от красных колбочек, а периферия — тормозящие сигналы от зеленых колбочек.
Следующий шаг состоял в том, чтобы рассмотреть рецептивные поля этих клеток, используя вместо рассеянного света маленькие цветные пятнышки; это мы и сделали в 1966 году совместно с Торстеном Визелом. У рецептивных полей большинства оппонентно-цветовых клеток де Валуа обнаружилась удивительная организация, которая до сих пор ставит нас в тупик. Клетки, как и у кошек по данным Куффлера, имели поля, разделенные на два антагонистических участка — центр и периферию; центр мог быть типа on или off. В типичном случае центр поля представлен исключительно красными, а тормозящая периферия — исключительно зелеными колбочками. Поэтому при красном свете как маленькое, так и большое пятно вызывает энергичную реакцию, поскольку центр избирательно чувствителен к длинноволновому свету, а периферия на него почти не реагирует; при коротковолновом свете маленькие пятна дают лишь очень слабую реакцию или не вызывают ее вовсе, а большие порождают сильное торможение с off-реакциями. При белом свете, содержащем короткие и длинные волны, малые пятна вызывают on-реакции, а большие не вызывают ответа. Хотя нашим первым впечатлением было, что такая клетка из центра должна получать входные сигналы от красных, а с периферии — от зеленых колбочек, теперь кажется вероятным, что полное рецептивное поле включает две перекрывающиеся системы, как это показано на рис. 127. Как красные, так и зеленые колбочки распределены по довольно широкой круговой области, причем их количество максимально в центре и убывает по мере удаления от него. В центре красные колбочки сильно преобладают, но к периферии число их падает значительно быстрее числа зеленых колбочек. Поэтому небольшое пятнышко, вспыхивающее в центре и содержащее длинноволновую компоненту, будет весьма мощным стимулом для красной системы; даже если оно стимулирует и зеленые колбочки, их число по сравнению с общим числом связанных с клеткой зеленых колбочек будет слишком мало, чтобы составить какую-либо конкуренцию красной системе. Те же соображения приложимы и к клеткам с центром и периферией, описанным в главе 3, рецептивные поля которых тоже должны состоять из двух оппонентных круговых перекрывающихся зон с разными формами кривых зависимости чувствительности от координаты. Таким образом, периферия, вероятно, имеет не кольцеобразную, как думали вначале, а круговую форму. В отношении этих оппонентно-цветовых клеток обезьян предполагается (правда, без достаточных данных), что периферические области отражают вклад горизонтальных клеток.
 Рис. 127. Графики вверху отражают зависимость чувствительности нейрона (измеренной, например, по реакции на неподвижное очень маленькое световое пятнышко) от положения стимула на сетчатке вдоль линии AA, проходящей через центр рецептивного поля. Для клетки с r-центром и g–-периферией красное пятнышко дает островершинную кривую, а зеленое — значительно более широкую кривую. Нижний график отображает реакцию на белый или желтый свет, стимулирующий обе оппонентные системы, так что происходит вычитание вкладов двух систем. При этом красные колбочки доминируют в центре, что приводит здесь к on-реакциям, а зеленые — на периферии, что приводит к off-реакциям.
Рис. 127. Графики вверху отражают зависимость чувствительности нейрона (измеренной, например, по реакции на неподвижное очень маленькое световое пятнышко) от положения стимула на сетчатке вдоль линии AA, проходящей через центр рецептивного поля. Для клетки с r-центром и g–-периферией красное пятнышко дает островершинную кривую, а зеленое — значительно более широкую кривую. Нижний график отображает реакцию на белый или желтый свет, стимулирующий обе оппонентные системы, так что происходит вычитание вкладов двух систем. При этом красные колбочки доминируют в центре, что приводит здесь к on-реакциям, а зеленые — на периферии, что приводит к off-реакциям.
Реакция на диффузный свет — в данном случае on-реакции на красный, off-реакции на синий и зеленый и отсутствие реакции на белый свет — ясно показывают, что такая клетка должна регистрировать информацию о цвете. Но реакции на определенного рода белые края и отсутствие реакций на диффузный свет убеждают в том, что эта клетка связана также и с восприятием формы. Мы называем эти цветооппонентные клетки с центром и периферией клетками «типа 1». Наружное коленчатое тело обезьяны, если вспомнить сведения из главы 4, содержит шесть слоев, причем четыре верхних слоя содержат множество мелких клеток, а два нижних — меньшее число крупных клеток. Мы находим клетки описанного выше типа 1 в верхних, или мелкоклеточных, слоях. Они различаются по типу колбочек, входящих в их центральную и периферийную системы, и по характеру центра, который может быть возбуждающим или тормозящим. Пример, представленный на рис. 126, мы можем обозначить как r+g– (с красным центром и зеленой периферией). Среди подтипов клеток, получающих входы от красных и зеленых колбочек, мы находим все четыре возможных варианта: r+ g–, r– g+, g+ r–, g– r+. Вторая группа клеток получает входы от синих колбочек из центра поля и от комбинации красных и зеленых (или, быть может, только от зеленых) — с периферии. Мы называем такие клетки «сине-желтыми», причем слово «желтые» здесь употреблено для краткости вместо «красные плюс зеленые». В четырех верхних слоях мы находим клетки еще двух типов. Клетки типа 2 составляют около 10 процентов нейронной популяции и имеют рецептивные поля, состоящие только из центра. Повсюду в этом центре у некоторых клеток выявляется красно-зеленая, а у остальных — сине-желтая оппонентность. Рецептивные поля еще примерно у 15 процентов клеток в четырех верхних слоях коленчатого тела и у всех клеток в двух нижних (крупноклеточных) слоях имеют центр и периферию, но эти клетки не проявляют цветовых предпочтений; создается впечатление, что центр и периферия их полей получают одинаковый относительный вклад от всех трех видов колбочек. Мы считаем эти клетки широкополосными и в верхних слоях называем их клетками типа 3. Все эти данные поразительно согласуются с моделью Геринга: кроме двух классов клеток с цветовой оппонентностью мы имеем также третий класс, вовсе не обладающий этим свойством, но с широкополосной пространственной оппонентностью. Что как будто не согласуется ни с какой теорией, так это пространственная организация полей у оппонентно-цветовых клеток — клеток типа 1. На первый взгляд можно подумать, что эта организация каким-то образом связана с цветовым контрастом, т.е. с тенденцией одного цвета, например синего, казаться более ярким в окружении другого цвета, например зеленого, или с тем, что кусок серой бумаги кажется желтоватым, если мы видим его на синем фоне. Но минутное размышление убедит вас, что клетки коленчатого тела едва ли могут быть полезны для этого вида цветового контраста: описанная выше клетка с r+-центром и g–-периферией не будет сильно возбуждена красным пятном на зеленом фоне — она ответит очень слабо или совсем не ответит, так как один эффект уничтожится другим; произойдет обратное тому, что нужно для усиления цветового контраста. О клетках типа 1 можно лишь утверждать, что по своей численности они должны быть наиболее важным для мозга источником цветовой информации, хотя эта информация представлена каким-то странным образом. Их возможная работа совместно с клетками типа 3 очень хорошо согласуется с представлениями Геринга о двух оппонентных цветовых системах и одной оппонентной пространственной системы. Вероятно, они играют также важную роль в восприятии тонких деталей формы объектов, поскольку единственные другие клетки коленчатого тела с маленькими центрами полей — это широкополосные клетки типа 3, которых в десять раз меньше. Как мы видели в главе 6, остроту зрения, т.е. способность различать малые объекты, можно измерять несколькими способами; можно, например, определять наименьшее расстояние между двумя точками, при котором они еще не сливаются, или наименьший видимый разрыв в окружности (так называемом кольце Ландольта). Острота, измеренная по любому из этих способов, оказывается для центральной ямки порядка 0,5 угловой минуты, или около 1 миллиметра с расстояния 8 метров. Этот показатель хорошо согласуется с расстоянием между двумя колбочками в центральной ямке. Клетки коленчатого тела типа 1, получающие входные сигналы от зоны центральной ямки, имеют рецептивные поля с центрами порядка 2 угловых минут в диаметре. Кажется вероятным, что центр поля составляет здесь всего одна колбочка. Таким образом, мы находим разумное соответствие между остротой зрения и наименьшими размерами центра рецептивного поля у клеток наружного коленчатого тела. Вентральная (нижняя) пара слоев коленчатого тела отличается от четырех дорсальных слоев тем, что состоит исключительно из клеток с широкополосными центрами полей. Эти клетки проявляют любопытную форму цветовой оппонентности, смысл которой никто не понимает и о которой я не буду говорить подробнее. Большинство считает эти клетки «слепыми» к цвету. Центры их полей в несколько раз крупнее, чем центры нейронов мелкоклеточных слоев, и у них есть ряд других интересных особенностей. В настоящее время мы подозреваем, что эти клетки обслуживают отделы мозга, играющие важную роль в восприятии формы, глубины и движения. Развитие этой темы увело бы нас далеко в сторону от цвета, и потребовалось бы написать еще одну книгу. Большинство типов клеток, описанных для наружного коленчатого тела, встречается также и в сетчатке. В коленчатом теле они больше обособлены, что облегчает их изучение. Вклад коленчатого тела в анализ зрительной информации у обезьян до сих пор не известен.
Нейронная основа константности цвета
Поскольку клетки типа 1 наружного коленчатого тела как будто не приспособлены для цвето-пространственных сравнений, нам, по-видимому, следует выйти на более высокие уровни зрительной системы. Для проверки идеи, согласно которой такие сравнения могли бы происходить в коре головного мозга, группа Лэнда, Маргарет Ливингстон и я изучали человека, у которого была произведена перерезка мозолистого тела с целью лечения эпилепсии. Между областями, разделенными средней вертикалью поля зрения, пространственно-цветовые взаимодействия у него отсутствовали: на цвет пятна, расположенного чуть левее точки, на которую смотрел этот испытуемый, не влияли даже резкие цветовые изменения в правом зрительном поле, тогда как нормальные испытуемые сообщали в таких случаях о заметных изменениях. Это означает, что цвето-пространственные взаимодействия, по-видимому, не могут происходить в самой сетчатке. Хотя никто всерьез и не утверждал обратного, вопрос продолжал дискутироваться, и было приятно получить какие-то экспериментальные данные. Результаты этого эксперимента согласуются с тем фактом, что нам не удалось обнаружить в сетчатке такие ганглиозные клетки, которые вполне могли бы участвовать в цвето-пространственных взаимодействиях. У золотой рыбки, осуществляющей пространственные сравнения не хуже нас с вами, коры головного мозга практически нет. Возможно, рыбка в отличие от нас выполняет эти сравнения с помощью своей сетчатки. Открытие в сетчатке рыб двойных оппонентных клеток (Н. Доу, 1968) как будто подтверждает это. У обезьян (см. следующий раздел) мы находим такие клетки в коре, но их нет ни в сетчатке, ни в наружном коленчатом теле.
Пузырьки
Примерно к 1978 году первичная зрительная кора обезьян с ее простыми и сложными клетками, с нейронами, реагирующими на концы линий, с глазодоминантными и ориентационными колонками представлялась достаточно изученной. Но неожиданной особенностью ее физиологии казалось то, что как будто лишь немногие клетки здесь небезразличны к цвету. Когда мы картировали рецептивное поле простой или сложной клетки, используя белый свет, а затем повторяли опыт с окрашенными пятнами или полосками, то результаты, как правило, были одинаковыми. Однако у некоторых клеток, составляющих, наверное, лишь десятую долю всех нейронов верхнего слоя коры, обнаруживались явные цветовые предпочтения — энергичные ответы на ориентированную полоску, скажем, красного цвета при полном отсутствии ответов на другие цвета или даже на белый цвет. Ориентационная избирательность этих клеток была не ниже, чем у клеток, нечувствительных к цвету, но большинство нейронов к цвету были безразличны. Все это казалось особенно странным ввиду того, что в наружном коленчатом теле очень большая доля клеток кодирует цвет, а коленчатое тело служит главным источником информации для зрительной коры. Трудно было понять, что могло происходить с этой цветовой информацией в коре. Неожиданно в 1978 году все изменилось. Нейроанатом из Сиэтла Маргарет Уонг-Райли обнаружила, что при окрашивании коры на фермент цитохромоксидазу в верхних слоях выявляется никогда не наблюдавшаяся ранее неоднородность — периодические темноокрашенные участки шириной около четверти миллиметра, разделенные промежутками около полумиллиметра. Цитохромоксидазу — участвующий в метаболизме фермент — содержат все клетки, и никто не мог даже подумать, что гистохимическая реакция на этот фермент позволит увидеть в коре что-либо интересное. Когда Уонг-Райли прислала нам свои микрофотографии, Торстен Визел и я заподозрили, что мы видим колонки глазодоминантности в поперечном разрезе и что большинство монокулярных клеток по каким-то причинам метаболически более активны, чем бинокулярные. Мы положили снимки в ящик стола и постарались забыть о них. Прошло несколько лет, прежде чем нам или кому-либо другому довелось изучать с помощью той же реакции срезы первичной зрительной коры, параллельные ее поверхности. Когда это приблизительно одновременно было, наконец, сделано двумя группами (Анитой Хендриксон и А. Хамфри в Сиэтле и Дж. Хортоном и мной в Бостоне), выявился — ко всеобщему полному изумлению — узор, напоминающий материал в горошек. Пример показан на рис. 129. Вместо полосок мы увидели похожие на пузырьки образования, которые ни с чем из известного ранее не связывались. Неоднородности Уонг-Райли называли всеми мыслимыми именами: точки, пуфы, мушки, пятнышки. Мы назвали их «пузырьками» [blobs — это слово наглядно, узаконено (оно есть даже в оксфордском словаре английского языка) и как будто раздражает наших конкурентов].
 Рис. 128. На этом поперечном срезе стриарной коры видны слои, окрашенные для выявления фермента цитохромоксидазы. Более темные области в слоях 2 и 3 (вертикальные полоски вверху) — «пузырьки».
Рис. 128. На этом поперечном срезе стриарной коры видны слои, окрашенные для выявления фермента цитохромоксидазы. Более темные области в слоях 2 и 3 (вертикальные полоски вверху) — «пузырьки».
 Рис. 129. Темные пятна представляют собой пузырьки, видимые «в плане»; около 50 таких пузырьков образуют характерный узор. Срез сделан через слой 3 параллельно поверхности коры на глубине около 0,5 мм. Срез проходит по границе между полями 17 (левая и средняя части) и 18 (правая часть), где пузырьков нет. (Желтые кружки — поперечные сечения кровеносных сосудов.)
Рис. 129. Темные пятна представляют собой пузырьки, видимые «в плане»; около 50 таких пузырьков образуют характерный узор. Срез сделан через слой 3 параллельно поверхности коры на глубине около 0,5 мм. Срез проходит по границе между полями 17 (левая и средняя части) и 18 (правая часть), где пузырьков нет. (Желтые кружки — поперечные сечения кровеносных сосудов.)
Следующая задача была очевидной: мы должны были снова регистрировать ответы клеток в стриарной коре, гистологически контролируя эксперимент окраской на цитохромоксидазу, и попытаться выявить нечто особенное у клеток, находящихся в пузырьках. В 1981 году Маргарет Ливингстон и я принялись за эту работу. Результат оказался совершенно неожиданным. Проходя расстояние в четверть миллиметра, равное диаметру пузырька, можно исследовать примерно пять или шесть клеток. Всякий раз, когда мы пересекали пузырек, попадавшиеся на пути электрода клетки были полностью лишены ориентационной избирательности, что заметно контрастировало с высокой ориентационной избирательностью клеток, расположенных вне пузырьков. Этому отсутствию ориентационной специфичности можно было бы дать два объяснения. Во-первых, эти клетки могли неизбирательно получать входные сигналы от соседних ориентационных клеток, лежащих вне пузырьков, и поэтому еще способны были отвечать специфической реакцией на линии (полоски и т.п.), но после объединения всех возможных ориентаций всякое предпочтение какой-либо из них исчезало. Во-вторых, они могли быть похожими на клетки коленчатого тела или на корковые клетки слоя 4C и, таким образом, быть проще внепузырьковых ориентационно-избирательных клеток. Вскоре вопрос был разрешен: оказалось, что большинство этих клеток имеет рецептивные поля с центром и периферией. Несколько дополнительных экспериментов убедили нас в том, что многие из них участвуют в кодировании цвета. Больше половины пузырьковых клеток имели оппонентно-цветовые рецептивные поля с центром и периферией, но вели себя явно более сложным образом, чем клетки типа 1 наружного коленчатого тела. Они практически не отвечали на белые пятна любых размеров и формы. Но на небольшие цветные пятна, вспыхивающие в центре рецептивного поля, они энергично реагировали в одном диапазоне длин волны и затормаживались в другом диапазоне; некоторые активировались длинными волнами (красным светом) и подавлялись короткими (зеленым и синим светом), другие вели себя противоположным образом. Как и среди клеток коленчатого тела, мы могли, в зависимости от положения максимальных реакций на спектральной шкале, выделить два класса — красно-зеленые и сине-желтые клетки. (Здесь, как и ранее, слова «красный», «зеленый» и «синий» указывают на соответствующие типы колбочек, а слово «желтый» — на параллельные входы от красных и зеленых колбочек.) Таким образом, эти клетки весьма напоминали оппонентно-цветовые клетки коленчатого тела, обладающие только центром (тип 2). Но в отличие от клеток типа 2 эти цветокодирующие пузырьковые клетки чаще всего не отвечали на большие белые или цветные пятна, каков бы ни был спектральный состав света. Они вели себя так, как если бы центральная система рецептивного поля была у них окружена кольцом оппонентности. Если говорить о наиболее распространенном типе клеток, то центр типа r+g–, казалось, был окружен у них кольцом типа r–g+. Мы с Маргарет Ливингстон назвали эти клетки двойными оппонентными из-за их красно-зеленой или желто-синей оппонентности в центре и антагонизма периферии по отношению к любой реакции в центре, будь она типа on или off. Поэтому они не отвечают не только на белый свет в любой геометрической конфигурации, но и на большие пятна, независимо от их спектрального состава. Центры рецептивных полей, как и у клеток типа 2, были в несколько раз больше, чем у клеток коленчатого тела типа 1. Как уже упоминалось, Н. Доу ввел термин двойные оппонентные для клеток, обнаруженных им в сетчатке золотой рыбки. Он предполагал, что сходные клетки могли быучаствовать в цвето-пространственных взаимодействиях у человека, и несколько лет спустя совместно с А. Пирлменом он старательно, хотя и безуспешно, искал такие клетки в наружном коленчатом теле макака. В конце 60-х годов и позже двойные оппонентные клетки изредка находили в коре обезьян, но их не связывали четко с какими-либо анатомическими структурами. Нам до сих пор непонятны некоторые из их особенностей. Например, у только что описанных клеток типа r+g– красное пятно, окруженное зеленым, часто вызывает слабую реакцию вместо энергичной, которой можно было бы ожидать. Вперемешку с двойными оппонентными клетками обоих классов (красно-зелеными и желто-синими) встречались и обычные широкополосные клетки с центром и периферией. Эти широкополосные клетки отличались от клеток верхних слоев коленчатого тела и от клеток слоя 4Cβ коры более крупными размерами своих центров. Мы с Маргарет Ливингстон предположили, что пузырьки представляют собой ветвь зрительного пути, имеющую дело с «цветом» в широком смысле, включая оттенки черного, белого и серого. Эта система, видимо, обособляется от остальной части зрительного пути либо в наружном коленчатом теле, либо в слое 4 стриарной коры. Коленчатое тело, возможно, имеет прямую, хотя и слабую проекцию на пузырьки. Кажется вероятной и проекция на них слоя 4Cβ — может быть, для них это даже главный источник входных сигналов. Проецируется ли на них слой 4Cα, неясно. Обычно для реакции пузырьковых клеток необходим граничный контраст — нужны либо границы яркости, на которые отвечают широкополосные клетки с центром и периферией, либо цветоконтрастные границы, на которые реагируют двойные оппонентные клетки. Как уже говорилось, это равнозначно тому, что такие клетки играют роль в создании константности цвета. Если пузырьковые клетки участвуют в константности цвета, они не могут осуществлять в точности те вычисления, которые рассматривались Лэндом, а именно сравнения между каким-то участком и его окружением для каждого из колбочковых спектральных поддиапазонов. Вместо этого они, по-видимому, осуществляют сравнение по типу Геринга: сопоставляют красно-зеленое в одном участке с красно-зеленым в окружающем фоне и делают то же самое для желто-синего и для интенсивности света. Но оба способа трактовки цвета — r, g и b в одном случае и b-w, r-g, и y-b в другом — по существу, эквивалентны. Цвет требует задания трех переменных: любому цвету соответствует тройка чисел, и мы можем представлять любой цвет точкой в трехмерном пространстве. Точки в таком пространстве можно представлять не единственным способом. Система координат может быть декартовой с осями, расположенными по нашему усмотрению, либо мы можем использовать полярные или цилиндрические координаты. В теории Геринга (в сетчатке и мозгу, наверное, тоже) просто используется иная совокупность осей для описания того же пространства. Эта теория, несомненно, упрощает действительность, так как пузырьковые клетки, принадлежащие к трем классам, вовсе не так сходны между собой, как горошины в стручке, — они значительно варьируют по относительной силе периферии и центра, по совершенству баланса между оппонентными цветами и по другим характеристикам, отчасти еще не вполне понятым. Сейчас мы можем лишь сказать, что физиология удивительно хорошо согласуется с психофизикой.
 Рис. 130. Вверху: согласно формулировке Лэнда, константность цвета обусловлена наличием трех типов клеток, сравнивающих активацию данного набора колбочек (красных, зеленых или синих) в какой-то области сетчатки со средней активацией такого же набора в окружающей зоне. Результат — три числа, определяющие цвет данной области. Например, желтому, коричневому, темно-серому и оливковому соответствуют определенные тройки чисел. Мы можем, таким образом, отображать цвета в некотором цветовом пространстве с тремя осями, которые соответствуют красному, зеленому и синему.
Внизу: математически эквивалентная система, в которой тоже могут быть заданы три числа. Она, вероятно, ближе к тому способу, которым мозг определяет цвета. В любой точке сетчатки мы можем говорить о степени «красно-зелености», измеряемой неким прибором, регистрирующим относительную силу стимуляции красных и зеленых колбочек (и показывающим нуль для желтого и белого). Эта величина определяется для данной области, а ее среднее значение — для окружения; затем вычисляется отношение этих величин. Тот же процесс повторяется для желто-синей и черно-белой систем. Получаемые три числа достаточны для задания любого цвета.
Рис. 130. Вверху: согласно формулировке Лэнда, константность цвета обусловлена наличием трех типов клеток, сравнивающих активацию данного набора колбочек (красных, зеленых или синих) в какой-то области сетчатки со средней активацией такого же набора в окружающей зоне. Результат — три числа, определяющие цвет данной области. Например, желтому, коричневому, темно-серому и оливковому соответствуют определенные тройки чисел. Мы можем, таким образом, отображать цвета в некотором цветовом пространстве с тремя осями, которые соответствуют красному, зеленому и синему.
Внизу: математически эквивалентная система, в которой тоже могут быть заданы три числа. Она, вероятно, ближе к тому способу, которым мозг определяет цвета. В любой точке сетчатки мы можем говорить о степени «красно-зелености», измеряемой неким прибором, регистрирующим относительную силу стимуляции красных и зеленых колбочек (и показывающим нуль для желтого и белого). Эта величина определяется для данной области, а ее среднее значение — для окружения; затем вычисляется отношение этих величин. Тот же процесс повторяется для желто-синей и черно-белой систем. Получаемые три числа достаточны для задания любого цвета.
Может возникнуть вопрос: зачем мозгу понадобились для отображения цвета такие на первый взгляд странные оси вместо более простой системы осей r, g и b, используемой в рецепторном слое сетчатки? По-видимому, цветовое зрение добавилось в ходе эволюции к «бесцветному» восприятию низших млекопитающих. Цветовое пространство у этих животных было одномерным, а колбочки разных типов (если животное обладало не одним типом колбочек) были объединены в общий пул. В ходе эволюции цветового зрения к уже существующей оси добавились еще две. Это было разумнее, чем отбросить уже сложившуюся черно-белую систему, а потом создавать три новые системы. Когда мы адаптируемся к темноте и используем только палочки, наше зрение лишается цвета и вновь отображается на единственную ось, в создании которой, очевидно, участвуют палочки. Это было бы не так просто сделать с осями r, g и b. В настоящее время мы можем лишь гадать, какова схема связей двойных оппонентных клеток. На рис. 131 показан один из возможных вариантов их соединения с клетками верхних слоев коленчатого тела или с клетками слоя 4Cβ коры. Излишне говорить, что все это чисто умозрительно; существенно лишь то, что двойная оппонентность каким-то образом должна достигаться: либо ее источник находится на нижних уровнях (сетчатки или коленчатое тело), и в этом случае мы просто еще не обнаружили клеток этого типа, либо это должно происходить в коре. Наличие таких клеток в сетчатке рыб не доказывает их существования у млекопитающих. Схема на рис. 131 всего лишь показывает, как могли бы быть сформированы центры рецептивных полей: для периферии мы всегда можем представить себе обратную процедуру с использованием клеток r–g+ коленчатого тела, поля которых широко перекрываются. Наше стремление рассматривать цвет и форму как раздельные аспекты восприятия находит, таким образом, подтверждение в виде физического обособления пузырьковых и непузырьковых участков первичной зрительной коры. В областях выше стриарной коры — в зрительной зоне 2 и даже еще выше — эта обособленность сохраняется. Мы не знаем, где происходит их объединение, если оно вообще где-нибудь происходит.
 Рис. 131. Двойная оппонентная клетка может быть построена с помощью многих клеток мелкоклеточного слоя наружного коленчатого тела. Большая окружность на рисунке очерчивает центр рецептивного поля двойной оппонентной клетки; если клетка принадлежит к типу r+g–, то ее входами могут быть многочисленные клетки с центром r+ и периферией g– и с меньшей величиной центральной части поля. Аналогично, периферия у двойной оппонентной клетки может складываться из элементов типа r–g+.
Рис. 131. Двойная оппонентная клетка может быть построена с помощью многих клеток мелкоклеточного слоя наружного коленчатого тела. Большая окружность на рисунке очерчивает центр рецептивного поля двойной оппонентной клетки; если клетка принадлежит к типу r+g–, то ее входами могут быть многочисленные клетки с центром r+ и периферией g– и с меньшей величиной центральной части поля. Аналогично, периферия у двойной оппонентной клетки может складываться из элементов типа r–g+.
Заключение
Самое интересное в цветовом зрении то, что здесь удается, объединив психофизические и нейрофизиологические методы, понять такие явления, как результаты смешения цветов или константность цвета, которые могли бы представляться совершенно загадочными. Проблемы, связанные с цветом, при всей их сложности, вероятно, проще тех, которые связаны с формой. Несмотря на открытие ориентационно-специфических клеток или клеток, реагирующих на концы линий, мы еще далеки от понимания нашей способности распознавать формы, отличать формы от их фона или воссоздавать трехмерную картину по плоским изображениям на сетчатке каждого глаза. Сопоставление этих двух модальностей — цвета и формы — вообще может ввести в заблуждение: вспомним, что одни лишь различия цвета на границах в отсутствие каких-либо различий освещенности могут приводить к восприятию формы. Цвет в этом смысле ничем не отличается от черного и белого и служит лишь одним из механизмов проявления формы.
9. Депривация и развитие
До сих пор мы представляли себе мозг как некий полностью сформированный, зрелый механизм. Нас интересовало то, каким образом соединены между собой его части, как эти части функционируют в каждодневных ситуациях и как они обслуживают интересы животного. Все это, однако, оставляло открытым совершенно иной и чрезвычайно важный вопрос: как этот механизм создается? В этой проблеме есть две главные составляющие. Развитие мозга в значительной части происходит до рождения животного, в материнской утробе. На первый взгляд мозг новорожденного младенца, хотя у него меньше складок и он несколько меньше мозга взрослого человека, в остальном не слишком от него отличается. Первый поверхностный взгляд, однако, вряд ли позволит выявить все подробности; ведь ребенок, понятно, не рождается со знанием алфавита или со способностью играть в теннис (или на арфе). Все эти достижения предполагают обучение, а под обучением мы, разумеется, понимаем формирование или модификацию нейронных связей под влиянием внешней среды. Таким образом, конечное состояние мозга — это результат как пренатального, так и постнатального развития. Во-первых, это развитие включает созревание как таковое, определяемое внутренними свойствами организма и происходящее до и после момента рождения; во-вторых, оно предполагает постнатальное созревание, определяемое тренировкой, обучением, образованием и опытом — все эти слова более или менее равнозначны. Пренатальное развитие — тема гигантская; мне известно о нем не слишком много, и поэтому я не буду описывать его сколько-нибудь подробно. Один из самых интересных, но и самых трудных вопросов состоит в том, каким образом отдельные нервные волокна огромного пучка отыскивают место своего назначения. Глаз, коленчатое тело и кора, например, формируются независимо друг от друга; по мере их созревания растущие из них аксоны должны делать выбор из множества альтернатив. Волокно зрительного нерва должно прорасти через сетчатку к слепому пятну, затем пройти в составе зрительного нерва к хиазме и принять здесь решение о том, следует ли переходить на противоположную сторону; затем оно должно проследовать к наружному коленчатому телу выбранной стороны, подойти к нужному слою (или к области, которая позднее превратится в нужный слой), а затем в точности к нужной части этого слоя так, что полученная в результате топография станет надлежащим образом упорядоченной; и наконец, оно должно разветвиться, причем веточки должны подойти к надлежащим частям клетки коленчатого тела — к ее телу или к дендриту. Сходные требования предъявляются к волокну, растущему от наружного коленчатого тела к зоне 17 или от зоны 17 к зоне 18. Хотя этот общий аспект развития нейронов в настоящее время интенсивно изучается во многих лабораториях, мы еще не знаем, каким образом волокна отыскивают места своего назначения. Трудно даже угадать победителя среди нескольких основных конкурирующих факторов, к которым относятся механическая наводка, химические градиенты и взаимодействие с комплементарными молекулами вроде происходящего в иммунной системе. Многочисленные современные исследования как будто бы указывают на роль многих различных механизмов. Рис. 132. По-видимому, щелевая форма зрачка, как у этой кошки и у многих ночных животных, позволяет более эффективно, чем круглый зрачок, снижать интенсивность света.
Рис. 132. По-видимому, щелевая форма зрачка, как у этой кошки и у многих ночных животных, позволяет более эффективно, чем круглый зрачок, снижать интенсивность света.
В этой главе мы рассмотрим в основном постнатальное развитие зрительной системы млекопитающих, особенно влияние, которое система испытывает со стороны окружающей среды. В отношении первых уровней зрительной системы кошки и обезьяны — сетчатки, коленчатого тела и, быть может, стриарной (первичной зрительной) коры — возникает естественный вопрос, сохраняется ли здесь после рождения некоторая пластичность. Я начну с описания простого эксперимента. Примерно к 1962 году стал известен ряд важных фактов о зрительной коре взрослой кошки: была открыта ориентационная избирательность, обнаружены «простые» и «сложные» клетки, о многих корковых нейронах стало известно, что они активируются сигналами от обоих глаз или проявляют различную степень глазодоминантности. Мы знали достаточно много о взрослом животном, чтобы задавать прямые вопросы относительно того, насколько пластична зрительная система. И вот Торстен Визел и я взяли котенка недельного возраста, когда глаза уже вот-вот должны открыться, и сшили его веки на одном глазу. Эта процедура может показаться жестокой, но она проводилась под наркозом, и проснувшийся котенок, возвращенный к своей матери и другим котятам, не выказал никаких признаков дискомфорта или страдания. Спустя десять недель мы хирургическим путем, опять-таки под наркозом, открыли глаз и стали регистрировать реакции корковых нейронов с целью выяснить, оказало ли отключение глаза какое-либо влияние на сам глаз или на зрительную систему. Прежде чем я опишу результаты, я должен отметить, что идея этого эксперимента была подсказана длительной историей психологических исследований и наблюдений, связанных с клинической неврологией. Психологи широко экспериментировали со зрительной депривацией на животных в 40-х и 50-х годах, используя для оценки результатов поведенческие методы. Типичный эксперимент состоял в том, что животное с момента рождения содержали в полной темноте. Когда животных выносили на свет, они оказывались слепыми или по меньшей мере обладали неполноценным зрением. Слепота была в некоторой степени обратимой, но зрительные функции восстанавливались лишь постепенно и обычно не полностью. Параллельно с этими экспериментами проводились клинические наблюдения над детьми с врожденной катарактой. Катаракта — это состояние, при котором хрусталик глаза мутнеет, он пропускает свет, но не позволяет формировать изображение на сетчатке. Катаракту у новорожденных, как и у взрослых, лечат путем хирургического удаления хрусталика и имплантации искусственной линзы или назначения очков с сильно выпуклыми стеклами. Это позволяет восстановить четкое изображение на сетчатке. Несмотря на относительную простоту операции, офтальмологи неохотно делали ее на очень маленьких детях или новорожденных, главным образом потому, что всякая операция, перенесенная в раннем детстве, статистически связана с большим риском, хотя этот риск и невелик. После удаления катаракты, скажем, в возрасте восьми лет и подбора очков результаты не оправдывали ожиданий. Зрение совсем не восстанавливалось: ребенок, как и раньше, оставался слепым, и глубокие дефекты сохранялись, несмотря на месяцы или годы попыток «научить его видеть». Ребенок, например, по-прежнему не мог отличить круг от треугольника. В результате появления и крушения надежд состояние ребенка вместо улучшения обычно ухудшалось. С этим резко контрастируют клинические случаи со взрослыми: у семидесятипятилетнего мужчины развивается катаракта на обоих глазах и он постепенно теряет зрение, но после трех лет слепоты катаракту удаляют, подбирают очки и зрение полностью восстанавливается. Зрение может даже улучшиться по сравнению с тем, что было до развития катаракты, поскольку хрусталик с возрастом желтеет и его удаление приводит к тому, что человек снова видит яркую синеву неба, которую воспринимают лишь дети и молодые люди. По-видимому, зрительная депривация (лишение зрительного опыта) у детей приводит к пагубным последствиям, никогда не наблюдаемым у взрослых. Обычно психологи вполне резонно объясняли результаты своих экспериментов, а также клинические результаты неспособностью ребенка «научиться видеть» или (что, по-видимому, то же самое) невозможностью образования связей из-за нехватки тренирующего опыта. Амблиопия — это частичная или полная потеря зрения, не связанная с какими-либо аномалиями самого глаза. Когда мы зашивали глаз у кошки или обезьяны, нашей целью было вызвать амблиопию, а затем выяснить, где в зрительном тракте возникнет аномалия. Результаты эксперимента с котенком поразили нас. Часто эксперимент дает неубедительные результаты, которые хороши лишь настолько, чтобы не опорочить идею полностью, но и не настолько определенны, чтобы вывести из них нечто полезное. Однако наш эксперимент явился исключением: результаты оказались совершенно ясными — когда мы открыли веки у котенка, сам глаз выглядел вполне нормальным; даже зрачок при его освещении сокращался обычным образом. Однако результаты регистрации активности корковых клеток никак нельзя было счесть нормальными. Хотя мы обнаружили много клеток с совершенно нормальными реакциями на ориентацию линий и их движение, мы нашли также, что вместо того, чтобы примерно половина клеток предпочитала один глаз другому, ни одна из двадцати пяти исследованных клеток не испытывала никакого влияния со стороны закрытого ранее глаза. (Пять клеток не испытывали влияния от обоих глаз, что изредка наблюдается даже у нормальных кошек.) Сравните это с нормальной кошкой, у которой около 15 % клеток монокулярны, причем около 7 % реагируют на левый глаз и 7 % — на правый. Гистограммы глазодоминантности для кошки, показанные вверху на рис. 133, позволяют сразу видеть это различие. Несомненно, что-то здесь основательно испортилось. Вскоре мы повторили этот эксперимент на других котятах и на детенышах обезьян. На котятах более обширная серия опытов вскоре показала, что при сшивании век сразу после рождения в среднем лишь 15 процентов клеток предпочитают зашитый ранее глаз против примерно 50 процентов у интактных котят. На обезьянах были получены примерно такие же результаты (см. нижние гистограммы на рис. 133). Среди немногочисленных клеток, отвечавших на стимуляцию закрытого ранее глаза, многие казались аномальными; они реагировали вяло, быстро утомлялись, и у них отсутствовала точная ориентационная настройка.
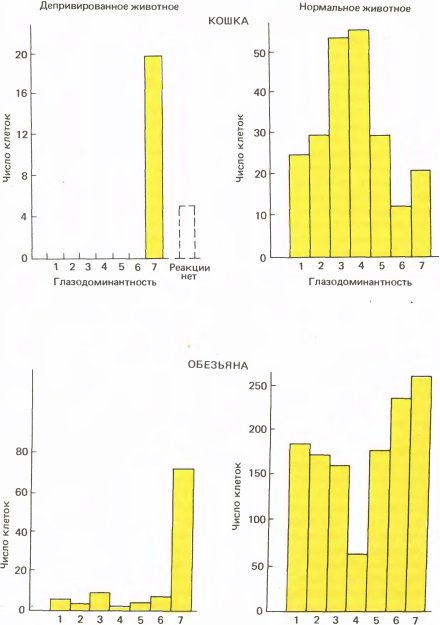 Рис. 133. Правый глаз котенка был на два с половиной месяца лишен зрительных стимулов в возрасте примерно десяти дней, когда глаза обычно уже открыты. В этом эксперименте (верхние гистограммы) мы зарегистрировали ответы только двадцати пяти клеток. (В последующих экспериментах мы смогли исследовать больше клеток и обнаружили, что небольшая доля их испытывала влияние со стороны ранее закрытого глаза.) В опытах на детеныше обезьяны (нижние гистограммы) результаты оказались весьма сходными. Его правый глаз закрыли в возрасте двух недель и держали закрытым на протяжении 18 месяцев. Позднее мы нашли, что те же результаты получаются и при закрытии глаза всего на несколько недель.
Рис. 133. Правый глаз котенка был на два с половиной месяца лишен зрительных стимулов в возрасте примерно десяти дней, когда глаза обычно уже открыты. В этом эксперименте (верхние гистограммы) мы зарегистрировали ответы только двадцати пяти клеток. (В последующих экспериментах мы смогли исследовать больше клеток и обнаружили, что небольшая доля их испытывала влияние со стороны ранее закрытого глаза.) В опытах на детеныше обезьяны (нижние гистограммы) результаты оказались весьма сходными. Его правый глаз закрыли в возрасте двух недель и держали закрытым на протяжении 18 месяцев. Позднее мы нашли, что те же результаты получаются и при закрытии глаза всего на несколько недель.
Результат, подобный этому, ставит множество вопросов. Где в зрительной системе возникала аномалия? В глазу? В коре? Могла ли кошка видеть закрытым ранее глазом, несмотря на корковую аномалию? Вызывалась ли эта аномалия просто световой депривацией или же лишением возможности видеть различные формы? Имел ли значение возраст, в котором мы закрывали глаз? Была ли аномалия результатом неиспользования глаза или она обусловливалась какими-то иными причинами? Чтобы все это выяснить, потребовалось много времени, но мы можем сформулировать результаты в нескольких словах. Для того чтобы определить локализацию дефекта, нужно было, очевидно, регистрировать реакции от более низких уровней, начиная, скажем, с сетчатки или с коленчатого тела. Результаты оказались однозначными: как в сетчатке, так и в коленчатом теле было множество клеток с практически нормальными реакциями. Клетки тех слоев коленчатого тела, к которым шли волокна от ранее закрытого глаза, имели обычные рецептивные поля с центром и периферией; они хорошо реагировали на маленькие пятна и плохо — на рассеянный свет. Единственным намеком на аномалию была некоторая вялость реакций этих клеток по сравнению с клетками слоев, получавших входные сигналы от нормального глаза. Зная об этой относительной нормальности, мы были поражены, впервые увидев под микроскопом наружное коленчатое тело, окрашенное по Нисслю. Его аномальность была видна практически и без микроскопа. Коленчатое тело у кошки организовано несколько проще, чем у обезьяны; оно состоит в основном из двух крупноклеточных слоев, расположенных наверху, а не внизу, как у обезьяны. Верхний слой имеет вход от контралатерального, а нижний — от ипсилатерального глаза. Под этими слоями лежит довольно плохо выраженный слой мелких клеток с несколькими подразделениями, которые я здесь игнорирую. На каждой стороне крупноклеточный слой, получавший входы от закрытого глаза, был бледнее и явно тоньше своего партнера, который выглядел совершенно нормальным. Клетки аномальных слоев были не только бледными, но и сморщенными примерно до двух третей своего нормального поперечного сечения. Результат, полученный при закрытии правого глаза, показан на рис. 134. Сходные результаты были получены и на макаках (рис. 135). Таким образом, мы столкнулись с парадоксом, на разрешение которого потребовалось несколько лет: в клетках наружного коленчатого тела, физиологически казавшихся сравнительно нормальными, гистологическое исследование показывало явную патологию. Как бы то ни было, мы теперь имели ответ на наш исходный вопрос, поскольку корковые клетки, практически не отвечавшие на стимуляцию ранее закрытого глаза, тем не менее, видимо, получали значительный и как будто бы нормальный поток входных сигналов от коленчатого тела; это позволяло думать, что первичное повреждение локализуется не в глазу или коленчатом теле, а главным образом в коре. При гистологическом исследовании коры мы не находили абсолютно никаких признаков аномалии. Как мы увидим далее, в коре все же были анатомические дефекты, но они не выявлялись с помощью применявшихся нами методов окраски.

 Рис. 134. Если правый глаз котенка закрыть в возрасте 10 дней на три с половиной месяца, то в его наружных коленчатых телах возникнут явные аномалии. В верхних частях обеих микрофотографий можно видеть два главных слоя. Вверху: верхний слой на левой стороне (контралатеральной по отношению к закрытому правому глазу) утончен и бледно окрашен. Внизу: на правой (ипсилатеральной) стороне аномален нижний из двух слоев. Оба слоя вместе имеют толщину около 1 мм. Гистограмма глазодоминантности показана на рис. 133.
Рис. 134. Если правый глаз котенка закрыть в возрасте 10 дней на три с половиной месяца, то в его наружных коленчатых телах возникнут явные аномалии. В верхних частях обеих микрофотографий можно видеть два главных слоя. Вверху: верхний слой на левой стороне (контралатеральной по отношению к закрытому правому глазу) утончен и бледно окрашен. Внизу: на правой (ипсилатеральной) стороне аномален нижний из двух слоев. Оба слоя вместе имеют толщину около 1 мм. Гистограмма глазодоминантности показана на рис. 133.
 Рис. 135. Поперечные срезы левого и правого наружных коленчатых тел обезьяны, правый глаз которой был закрыт в возрасте двух недель на 18 месяцев. Видны аномальные слои, они выглядят более бледными. На обеих сторонах это те слои, которые получают входные волокна ранее закрытого (правого) глаза,— слои 1, 4 и 6 слева, 2, 3 и 5 справа (счет идет снизу вверх).
Клетки измененных слоев мельче, но это невозможно рассмотреть при таком малом увеличении. Ширина всей структуры около 5 миллиметров.
Рис. 135. Поперечные срезы левого и правого наружных коленчатых тел обезьяны, правый глаз которой был закрыт в возрасте двух недель на 18 месяцев. Видны аномальные слои, они выглядят более бледными. На обеих сторонах это те слои, которые получают входные волокна ранее закрытого (правого) глаза,— слои 1, 4 и 6 слева, 2, 3 и 5 справа (счет идет снизу вверх).
Клетки измененных слоев мельче, но это невозможно рассмотреть при таком малом увеличении. Ширина всей структуры около 5 миллиметров.
Следующий вопрос заключался в том, что же именно порождает аномалию при закрытии глаза. Сшивание век уменьшает количество света, достигающего сетчатки, приблизительно в 10–50 раз; оно, разумеется, препятствует также созданию на сетчатке каких-либо изображений. Может ли аномалия вызываться просто уменьшением количества света? Чтобы выяснить это, мы вставляли в один глаз новорожденного котенка матовую контактную линзу из пластмассы, по консистенции близкой к материалу мячика для настольного тенниса. У других животных мы вместо этого пришивали на одном глазу тонкую просвечивающую мембрану, играющую роль дополнительного века и называемую мигательной перепонкой, которая имеется у кошек, но отсутствует у людей. Пластмасса или перепонка уменьшала количество света всего лишь вдвое, но предотвращала формирование каких-либо сфокусированных изображений. Результаты оказались такими же: аномальная корковая физиология и аномальная гистология коленчатого тела. Очевидно, повреждение вызывалось невозможностью воспринимать формы, а не просто недостатком света. У нескольких котят перед регистрацией нейронных ответов мы тестировали зрение, надевая непрозрачную черную контактную линзу на тот глаз, который не был закрыт, и затем наблюдая за поведением животного. Животные были, несомненно, слепы на депривированный глаз: будучи помещены на низком столике, они уверенно шли к его краю, перешагивали его и падали на разложенную на полу подстилку. Ходя по полу, они обычно натыкались на ножки стола. Этого никогда не делает ни одна нормальная, уважающая себя кошка. Аналогичные тесты с незакрытым глазом показали, что зрение было совершенно нормальным. Далее мы провели длительные исследования на кошках и обезьянах с целью выяснить, зависят ли результаты от возраста, в котором закрывают глаз, и от продолжительности депривации. Вскоре выяснилось, что возраст имеет решающее значение. У взрослой кошки, у которой один глаз был отключен более чем на год, не наблюдалось ни слепоты на этот глаз, ни потери корковых реакций, ни патологии коленчатого тела. (Первая депривированная кошка, мать нашего первого помета котят, была взрослой по определению!) После многочисленных экспериментов мы пришли к выводу, что где-то между рождением и зрелым возрастом должен существовать период пластичности, когда депривация приводит к корковому дефекту. Как выяснилось, у кошки этот критический период длится от четвертой недели до четвертого месяца. Неудивительно, что закрытие глаза мало влияет до наступления четвертой недели, так как в первый месяц жизни кошка почти не пользуется зрением: глаза открываются лишь около 10-го дня, а несколько последующих недель котята со своей матерью прячутся за диваном. Чувствительность к депривации быстро возрастает и достигает максимума в первые недели критического периода. В это время отключение одного глаза даже на несколько дней приводит к заметному искажению гистограммы глазодоминантности. В последующие четыре месяца время депривации, необходимое для получения явных эффектов, неуклонно возрастает; иными словами, чувствительность к депривации уменьшается и сходит на нет. На рис. 136 представлены гистограммы, иллюстрирующие некоторые результаты, полученные на обезьянах. На левом графике — эффект отключения в возрасте пяти дней одного глаза на 6 недель; почти нет клеток, которые отвечали бы на стимуляцию ранее закрытого глаза. Гораздо более кратковременная депривация (средний график) тоже вызывает сильно выраженный эффект, но явно меньший, чем при более длительной депривации. В возрасте четырех месяцев чувствительность снижается настолько, что даже отключение глаза на 5 лет (правый график) дает эффект хотя и весьма заметный, но все же несравнимый с последствиями более ранней депривации.

Рис. 136. Слева: у обезьяны, правый глаз которой был зашит в возрасте пяти дней на шесть недель, практически полностью доминирует левый глаз. В середине: закрыть глаз всего на несколько дней в возрасте нескольких недель уже достаточно, чтобы вызвать заметный сдвиг глазодоминантности. Более темным цветом показано число аномальных клеток. Справа: если закрытие глаза отложить до четырехмесячного возраста, то очень длительное его отключение (в данном случае на пять лет) приводит к гораздо меньшему сдвигу глазодоминантности, чем кратковременное отключение в возрасте нескольких недель.
При изучении периода чувствительности у кошек и обезьян были получены весьма сходные результаты. У обезьян этот период начинается раньше, в момент рождения, а не в возрасте четырех недель, и длится дольше, постепенно заканчиваясь к году, а не к четвертому месяцу. Чувствительность наиболее высока в первые две недели жизни, на протяжении которых всего нескольких дней депривации оказывается достаточным, чтобы вызвать выраженный сдвиг глазодоминантности. Отключение глаза у взрослой обезьяны независимо от его продолжительности не вызывает никаких вредных последствий. У одной взрослой обезьяны мы закрывали глаз на пять лет, после чего не отмечалось ни слепоты, ни коркового дефекта, ни сморщивания клеток в коленчатых телах.
Восстановление
Следующим интересовавшим нас вопросом был вопрос о возможности какого-либо восстановления нарушенной функции у обезьяны после открытия отключенного ранее глаза. Оказалось, что никакого или почти никакого физиологического восстановления не происходит, если глаз, закрытый на неделю или на больший срок, просто открыть и ничего более не делать. Даже спустя несколько лет кора оставалась приблизительно такой же аномальной, как и в момент открытия глаза (рис. 137, левый график). Если же, открыв один глаз, закрывали другой, ранее открытый (эту процедуру мы назвали реверсией), то восстановление наблюдалось, но лишь в том случае, если обезьяна все еще находилась в критическом периоде (рис. 137, средний и правый графики — для случая ранней и поздней реверсии). После окончания критического периода даже реверсия — закрытие другого глаза на несколько лет — не давала практически ничего, кроме незначительного восстановления структуры или функции. Способность обезьяны видеть не всегда соответствовала физиологическому состоянию зрительной коры. Без реверсии зрение в ранее закрытом глазу никогда не восстанавливалось. При реверсии зрение возвращалось и часто достигало почти нормального уровня, причем это происходило даже в случаях поздней реверсии, когда физиология первоначально закрытого глаза оставалась весьма аномальной. Мы все еще не можем объяснить это несоответствие между отсутствием существенной физиологической или анатомической нормализации и тем, что в ряде случаев зрение, по-видимому, в значительной степени восстанавливалось. Возможно, что те и другие наблюдения отражают разные стороны зрительной функции. Мы испытывали остроту зрения, измеряя такие показатели, как наименьший различимый разрыв в линии или окружности. Такое тестирование, однако, может не давать полного представления о зрительной функции. Трудно поверить, что столь выраженные физиологические и анатомические дефекты проявляются на поведенческом уровне всего лишь в незначительном снижении остроты зрения.
 Рис. 137. Слева: у макака один глаз закрыли при рождении на 9 дней, а затем открыли. Ответы клеток регистрировали по прошествии четырех лет, на протяжении которых зрение животного многократно тестировалось. Даже столь длительный период, когда оба глаза были открыты, не привел к заметному восстановлению физиологических показателей.
В середине: правый глаз был отключен с рождения; через пять с половиной недель этот глаз открыли, а левый закрыли. Когда в возрасте шести месяцев регистрировали ответы в правом полушарии, в отношении большинства клеток резко доминировал первоначально закрытый правый глаз.
Справа: в этом случае правый глаз закрыли в возрасте 7 дней на один год, после чего открыли его и закрыли левый глаз. Спустя еще год открыли и левый глаз, после чего оба глаза оставались открытыми. Когда, наконец, в возрасте трех с половиной лет регистрировали корковые ответы, большинство клеток реагировало на сигналы от того глаза, который был открыт в самом начале. Очевидно, один год — слишком поздний срок для реверсии.
Рис. 137. Слева: у макака один глаз закрыли при рождении на 9 дней, а затем открыли. Ответы клеток регистрировали по прошествии четырех лет, на протяжении которых зрение животного многократно тестировалось. Даже столь длительный период, когда оба глаза были открыты, не привел к заметному восстановлению физиологических показателей.
В середине: правый глаз был отключен с рождения; через пять с половиной недель этот глаз открыли, а левый закрыли. Когда в возрасте шести месяцев регистрировали ответы в правом полушарии, в отношении большинства клеток резко доминировал первоначально закрытый правый глаз.
Справа: в этом случае правый глаз закрыли в возрасте 7 дней на один год, после чего открыли его и закрыли левый глаз. Спустя еще год открыли и левый глаз, после чего оба глаза оставались открытыми. Когда, наконец, в возрасте трех с половиной лет регистрировали корковые ответы, большинство клеток реагировало на сигналы от того глаза, который был открыт в самом начале. Очевидно, один год — слишком поздний срок для реверсии.
Природа повреждений
Описанные выше результаты ясно показали, что отсутствие изображений на сетчатке в раннем периоде жизни ведет к глубоким и стойким нарушениям корковой функции. Оставались, однако, открытыми два важных вопроса относительно природы этого явления. Первым из них был вопрос, связанный с дилеммой «природа или воспитание»: лишали ли мы животных индивидуального опыта, в котором они нуждались для создания правильных связей, или же мы разрушали те связи, которые уже существовали, были заранее сформированы и функционировали при рождении животного? Практически все эксперименты с выращиванием животных в темноте, проведенные за десятки лет до нашей работы, интерпретировались в контексте проблемы обучаемости. Кору головного мозга, которую большинство людей считали (и продолжают считать) физиологическим субстратом памяти и мышления, рассматривали как нечто подобное запоминающему устройству емкостью в 1 мегабайт, за которое мы так дорого платим при покупке компьютера: и в том и в другом содержится множество элементов и соединений, но нет никакой информации, пока мы ее туда не заложим. Короче говоря, кору считали своего рода tabula rasa.
 Рис. 138. Японский макак (Macaca fuscata), самый крупный из всех макаков, обитает на земле и на деревьях в северной части Японии. Он защищен от холода густым серовато-бурым мехом.
Рис. 138. Японский макак (Macaca fuscata), самый крупный из всех макаков, обитает на земле и на деревьях в северной части Японии. Он защищен от холода густым серовато-бурым мехом.
Одним из очевидных подходов к решению поставленного вопроса могла бы быть прямая регистрация активности нейронов у новорожденных котят или обезьян. Если бы для создания нужных связей требовалось обучение, мы не обнаружили бы ничего похожего на ту высокоразвитую специфичность, которая наблюдается у взрослых животных. Отсутствие специфичности, однако, не решило бы проблему, так как неразвитость связей можно было бы объяснить либо незрелостью (тем, что построение генетически запрограммированной системы связей еще не завершилось), либо отсутствием сенсорного опыта. С другой стороны, обнаружение такой специфичности говорило бы против механизма обучения. Мы не ожидали, что эксперименты на котятах будут простыми; так оно и оказалось. Котенок рождается с очень незрелой зрительной системой и совсем не использует глаза примерно до десятого дня, когда они открываются. В это время даже среды глаза, прозрачные субстанции, находящиеся между роговицей и сетчаткой, еще не вполне прозрачны и не позволяют получить четкое изображение на сетчатке. Незрелая зрительная кора действительно реагировала вяло, несколько непредсказуемо и в целом сильно отличалась от нормальной зрительной коры взрослой кошки; тем не менее мы нашли много клеток с явной ориентационной специфичностью. Чем больше времени проходило от рождения до регистрации, тем большее число клеток по своему поведению приближалось к клеткам взрослого типа — может быть, из-за прояснения глазных сред и общего укрепления организма, а быть может, и в результате обучения. Различные группы исследователей объясняют это по-разному. Наиболее убедительные данные были получены на новорожденных обезьянах. У макака на другой день после рождения зрительная система обнаруживает поразительную зрелость: детеныши смотрят в разные стороны, следят взором за предметами и проявляют большой интерес к окружающему (рис. 139). В соответствии с таким поведением клетки первичной зрительной коры у них проявляют столь же острую ориентационную настройку, как и у взрослой особи. Последовательно расположенные клетки хорошо упорядочены по ориентации (см. график на рис. 140). Хотя мы и отмечали некоторые различия между новорожденными и взрослыми животными, система ориентационно-специфичных рецептивных полей, эта отличительная особенность стриарной коры, казалась вполне четко организованной.
 Рис. 139. На следующий день после рождения макак-детеныш осматривается, фиксирует взором предметы, проявляет живой интерес к окружающему миру. У человека и кошки зрительная функция достигает такой степени зрелости лишь спустя много недель после рождения.
Рис. 139. На следующий день после рождения макак-детеныш осматривается, фиксирует взором предметы, проявляет живой интерес к окружающему миру. У человека и кошки зрительная функция достигает такой степени зрелости лишь спустя много недель после рождения.
 Рис. 140. У новорожденного макака клетки коры проявляют примерно такую же ориентационную избирательность, как и у взрослых особей, и последовательность ориентаций почти так же упорядоченна.
Рис. 140. У новорожденного макака клетки коры проявляют примерно такую же ориентационную избирательность, как и у взрослых особей, и последовательность ориентаций почти так же упорядоченна.
У макака (в сравнении с кошкой или человеком) зрительная система новорожденной особи может считаться весьма зрелой, но она, несомненно, анатомически отличается от зрительной системы взрослой обезьяны. Срезы коры, окрашенные по Нисслю, выглядят различно: у детенышей более тонкие слои и более плотная упаковка клеток. Ле-Вэй впервые показал, что даже общая площадь стриарной коры от рождения к зрелому возрасту увеличивается примерно на 30 процентов. Если окрасить кору по методу Гольджи или исследовать ее под электронным микроскопом, отличия станут еще более очевидными: у клеток обычно менее развита система дендритов и меньше синапсов. Ввиду этих различий было бы удивительно, если бы кора уже при рождении функционировала в точности так же, как у взрослого животного. С другой стороны, через месяц после рождения дендриты и синаптические связи все еще не достигают полного развития. Вопрос «природа или воспитание?» означает: определяется ли постнатальное развитие опытом или все еще протекает по врожденной программе? Мы пока не уверены в ответе, но, судя по сравнительной нормальности реакций корковых клеток при рождении, можно сделать вывод, что нарушение их после депривации обусловлено в основном «порчей» связей, имевшихся к моменту рождения, а не невозможностью их формирования без индивидуального опыта. Второй важный вопрос касается причины этой порчи. На первый взгляд ответ казался почти очевидным. Мы полагали, что деградация связей обусловлена их неиспользованием, подобно тому как происходит атрофия мышц ноги при фиксации колена или лодыжки гипсовой повязкой. В сморщивании клеток коленчатого тела мы видели проявление постсинаптической атрофии — так называют уменьшение величины клеток в наружных коленчатых телах взрослых животных или людей после удаления глаза. Эти предположения оказались неверными. Мы считали их настолько самоочевидными, что я не уверен, что мы когда-либо подумали бы о проведении специального эксперимента для их проверки. Нам пришлось изменить свою точку зрения только в результате одного необязательного, как нам тогда казалось, эксперимента, причин постановки которого я уже не помню. Мы зашили оба глаза сначала у новорожденного котенка, а потом у новорожденного детеныша обезьяны. Если бы нарушение реактивности корковых клеток, получающих входные сигналы от одного глаза, было обусловлено их бездействием, то отключение обоих глаз удвоило бы дефект: мы практически не смогли бы обнаружить клеток, реагирующих на левый или правый глаз. На самом же деле, к нашему великому изумлению, мы не получили ничего похожего на отсутствие клеточных реакций: после открытия глаз добрая половина клеток стриарной коры снова отвечала нормально, одна четверть отвечала аномально и одна четверть не реагировала вовсе. Мы вынуждены были заключить, что невозможно предсказать судьбу корковой клетки в ситуации, когда закрыт один глаз, если неизвестно, был ли при этом закрыт и другой. Отключите один глаз, и клетка почти наверняка утратит свои связи с ним; но отключите оба глаза, и у вас будут неплохие шансы на то, что связь сохранится. Мы, очевидно, столкнулись здесь не с бездействием клеток и их связей, а с какого-то рода конкуренцией между двумя глазами. Дело обстоит так, как если бы клетка вначале имела две группы синаптических входов — два входных пути, по одному от каждого глаза, но при неиспользовании одного пути одерживает верх другой, завладевая территорией первого (рис. 141).
 Рис. 141. Мы предполагаем, что корковая клетка имеет входы от двух источников, по одному от каждого глаза, и что закрытие одного глаза ослабляет связи от этого глаза и усиливает связи от другого.
Рис. 141. Мы предполагаем, что корковая клетка имеет входы от двух источников, по одному от каждого глаза, и что закрытие одного глаза ослабляет связи от этого глаза и усиливает связи от другого.
Такое рассуждение, мы полагали, вряд ли применимо к сморщиванию клеток коленчатого тела, поскольку эти клетки монокулярны и здесь не видно каких-либо возможностей конкуренции. В то время мы не смогли объяснить атрофию клеток в слоях, соответствующих закрытому глазу. При закрытии обоих глаз сморщивание клеток коленчатого тела не столь заметно, но здесь трудно что-либо утверждать, так как при этом отсутствуют нормальные слои, которые можно было бы использовать для сравнения. Наше понимание всей этой проблемы не продвинулось, пока мы не начали использовать некоторые новые методы экспериментальной анатомии.
Косоглазие
Наиболее частой причиной амблиопии у людей бывает косоглазие, или страбизм — непараллельность оптических осей глаз. Косоглазие может быть сходящимся или расходящимся. Причина косоглазия неизвестна, почти наверняка существует несколько причин. Иногда косоглазие возникает после рождения, в первые месяцы жизни, когда глаза едва-едва начинают фиксировать и прослеживать предметы. Нарушения прямоты взгляда могут быть результатом аномалий глазных мышц или механизмов мозгового ствола, обслуживающих движение глаз. У некоторых детей косоглазие, по-видимому, связано с дальнозоркостью. Для правильной фокусировки удаленного объекта хрусталик дальнозоркого глаза должен принять столь же выпуклую форму, как хрусталик нормального глаза, когда он фокусирует близкий объект. Округление хрусталика при рассматривании близких предметов означает сокращение цилиарной мышцы внутри глаза; этот процесс называют аккомодацией. Когда человек с нормальным зрением осуществляет аккомодацию на близкое расстояние, глаза одновременно автоматически поворачиваются к срединной плоскости (конвергенция). Эти два процесса показаны на рис. 142. Нейронные сети мозгового ствола, управляющие аккомодацией и конвергенцией, вероятно, связаны друг с другом и, возможно, перекрываются; как бы то ни было, трудно осуществить одну из этих реакций без другой. Когда дальнозоркий человек осуществляет аккомодацию, которая необходима ему для фокусировки даже удаленного объекта, один или оба глаза могут повернуться внутрь, хотя конвергенция в этом случае нежелательна. Если дальнозоркий ребенок не носит очки, поворот глаза может стать со временем постоянной привычкой. Это объяснение косоглазия, конечно, справедливо лишь в некоторых, но не во всех случаях, поскольку косоглазие не всегда сопровождается дальнозоркостью, а у некоторых людей с косоглазием один глаз поворачивается кнаружи, а не вовнутрь. Косоглазие можно излечить хирургическим путем, изменив места прикреплениянаружных мышц глаза. Операция обычно с успехом исправляет положение глазных осей, но до последнего десятилетия ее чаще всего откладывали до тех пор, пока ребенок не достигнет возраста четырех — десяти лет (по той же причине, по которой откладывали удаление катаракты, — чтобы несколько уменьшить риск операции). Косоглазие, возникающее у взрослых, например вследствие повреждения нерва или глазной мышцы, сопровождается, конечно, двоением в глазах. Чтобы испытать это состояние, достаточно лишь слегка надавить на один глаз снизу и сбоку. Двоение в глазах может быть весьма неприятным и даже выводить человека из строя, и если никакое лучшее решение невозможно, один глаз следует закрыть повязкой. В противном случае двоение в глазах останется до тех пор, пока не будет исправлено косоглазие. У детей с косоглазием, однако, двоение в глазах редко продолжается длительное время; оно сменяется либо поочередным использованием двух глаз, либо подавлением зрения в одном глазу.
 Рис. 142. При рассматривании близкого предмета происходят две приспособительные реакции: хрусталик округляется в результате сокращения цилиарных мышц, и глаза поворачиваются к срединной плоскости головы.
Рис. 142. При рассматривании близкого предмета происходят две приспособительные реакции: хрусталик округляется в результате сокращения цилиарных мышц, и глаза поворачиваются к срединной плоскости головы.
Когда ребенок чередует глаза, он фиксирует предметы сначала одним глазом, в то время как нефиксирующий глаз поворачивается внутрь или наружу, а затем другим глазом, а первый при этом отводится в сторону. (Чередующееся косоглазие весьма распространено, и если вы знаете его признаки, вы легко распознаете его.) Глаза используются по очереди, иногда смена происходит почти ежесекундно, и пока один глаз смотрит, другой как бы ничего не видит. В любой момент, когда один глаз расположен правильно, а другой отклоняется, говорят, что зрение в отклоненном глазу подавлено. Подавление знакомо всякому, кто привык смотреть в монокулярный микроскоп, прицеливаться из ружья или выполнять какую-либо иную задачу, используя только один глаз при открытом другом глазе. Для подавленного глаза зрительное восприятие попросту исчезает. Ребенок с чередующимся косоглазием всегда подавляет тот или другой глаз, но если проверить зрение в каждом глазу по отдельности, то обычно окажется, что оба глаза вполне нормальны. Некоторые дети с косоглазием не чередуют глаза, а используют все время один глаз при подавленном другом. Если один глаз привычно подавляется, то зрение в подавляемом глазу проявляет тенденцию к ухудшению. Острота зрения падает, особенно в области центральной ямки и около нее, и, если это состояние длится достаточно долго, глаз может стать практически слепым. Такого рода слепоту офтальмологи называют amblyopia ex anopsia. Это наиболее распространенный вид амблиопии и даже слепоты вообще. Естественно, что нам пришло в голову попытаться вызвать косоглазие и, следовательно, амблиопию у котят или обезьяны путем хирургической перерезки одной из мышц глаза после рождения, так как при этом мы смогли бы провести физиологические исследования и посмотреть, какая часть зрительного пути нарушена. Мы сделали это на полудюжине котят и к своему неудовольствию обнаружили, что у котят, как и у многих детей, развилось чередующееся косоглазие; они смотрели сначала одним глазом, а потом другим. Протестировав каждый глаз в отдельности, мы вскоре убедились, что в обоих глазах зрение было нормальным. Стало ясно, что нам не удалось вызвать амблиопию, и мы начали думать, что же делать дальше. Мы решили провести на одном из котят регистрацию клеточных реакций, хотя совершенно не представляли себе, что можно было таким образом выяснить (исследования часто включают поиски «ощупью»). Результаты оказались совершенно неожиданными. Изучая клетку за клеткой, мы вскоре поняли, что с мозгом случилось нечто странное: каждая клетка реагировала совершенно нормально, но только на стимуляцию одного глаза. По мере продвижения электрода в коре шел ряд нейронов, активируемых левым глазом, затем этот ряд обрывался и в действие вступал другой глаз. В отличие от того, что мы наблюдали после закрытия глаза, ни один глаз не был ущемлен в своих правах по сравнению с другим в общем балансе доминирования. Иногда в зоне перехода появлялись бинокулярные клетки, но, как видно из рис. 143, поля бинокулярных клеток во всей популяции составляли около 20 процентов вместо 85 в норме.
 Рис. 143. Если у новорожденного котенка перерезать одну из наружных мышц глаза, спустя три месяца подавляющее большинство клеток оказываются монокулярными, попадая в группы 1 и 7.
Рис. 143. Если у новорожденного котенка перерезать одну из наружных мышц глаза, спустя три месяца подавляющее большинство клеток оказываются монокулярными, попадая в группы 1 и 7.
Мы заинтересовались тем, что произошло с исходно бинокулярными клетками — может быть, они просто погибли или совсем утратили способность реагировать, так что оставались только первично монокулярные клетки? Это казалось весьма маловероятным, так как по мере продвижения электрода мы находили обычное изобилие реагирующих клеток — не было ничего похожего на пятикратное уменьшение их общего числа. У нормальной кошки при горизонтальном продвижении электрода в верхних слоях коры мы обычно встречаем примерно от 10 до 15 клеток подряд с доминированием одного и того же глаза; все они явно принадлежат к одной и той же колонке глазодоминантности, и две или три из них могут быть монокулярными. У косоглазых котят мы точно так же наблюдали по 10–15 клеток с доминированием одного глаза, но теперь все, кроме двух или трех, были монокулярными. Видимо, каждая клетка полностью или почти полностью перешла в ведение того глаза, который ранее просто оказывал на нее большее влияние. Чтобы оценить значение этого результата, следует вспомнить, что мы в сущности никак не изменили общее количество зрительных стимулов, получаемых каждой сетчаткой. Поскольку у нас не было оснований полагать, что мы повредили какой-либо из двух глаз, мы пришли к выводу, что общий поток импульсов в двух зрительных нервах должен был остаться нормальным. Каким же образом косоглазие смогло вызвать столь радикальные изменения корковой функции? Чтобы ответить на этот вопрос, нам нужно рассмотреть, как два глаза взаимодействуют друг с другом в норме. Косоглазие изменило связь между стимулами, воздействующими на оба глаза. Когда мы рассматриваем какой-то объект, изображения любой его точки в норме попадают на те места двух сетчаток, которые находятся на одинаковом расстоянии и в одинаковом направлении относительно обеих центральных ямок, — они попадают на корреспондирующие точки. Если изображение на левой сетчатке активирует какую-то бинокулярную клетку (это происходит, если рецептивное поле клетки пересекается границей темнота/свет, ориентация которой в точности соответствует данной клетке), то эта клетка будет также активироваться изображением на правой сетчатке. Это обусловлено тремя причинами: 1) изображения попадают на одни и те же участки обеих сетчаток; 2) бинокулярная клетка (если только она не специализирована для восприятия глубины) имеет рецептивные поля точно в одних и тех же участках обеих сетчаток; 3) ориентационные предпочтения бинокулярных клеток всегда близки для обоих глаз. Очевидно, что в случае косоглазия причина 1 отпадает: если при несовпадающих изображениях один глаз в данный момент побуждает клетку разряжаться, то будет ли делать то же самое другой глаз, зависит только от случая. Для одиночной клетки это, по-видимому, единственный фактор, изменяющийся при косоглазии. Когда котенок несколько недель или месяцев находится в такой ситуации, при которой сигналы от обоих глаз более не соответствуют друг другу, это приводит к тому, что более слабая из двух групп входных связей данной клетки становится еще слабее и часто практически исчезает. Таким образом, мы имеем здесь пример патологических изменений, вызванных не отсутствием стимуляции, а всего лишь разрывом нормальных временны́х соотношений между двумя совокупностями стимулов, — изощренный удар, если учитывать серьезность последствий. В аналогичных экспериментах на обезьянах были получены такие же результаты; поэтому кажется, что косоглазие приводит к тем же последствиям и у людей. По клиническим данным, у человека с длительным чередующимся косоглазием даже после устранения этого дефекта обычно не восстанавливается способность к восприятию глубины. Хирург может привести оба глаза в соответствие лишь с точностью до нескольких градусов, а больной, вероятно, уже утратил способность достигать полного слияния двух изображений путем правильной установки глаз с точностью до нескольких угловых минут. Хирургическое исправление косоглазия выравнивает глаза достаточно хорошо, и нормальные нейронные механизмы справились бы с оставшимися несколькими градусами тонкой подстройки, но при косоглазии именно эти механизмы, включающие бинокулярные клетки коры, оказываются нарушенными. Для восстановления, вероятно, потребовался бы длительный период точного выравнивания глаз, который предполагает как нормальную мышечную подстройку, так и подстройку, определяемую бинокулярным зрением. Все эти соображения, объясняющие сдвиг глазодоминантности, весьма напоминают модель, позволяющую объяснить ассоциативное научение изменениями в синапсах. Эта модель была предложена психологом Дональдом Хеббом из Университета Мак-Гилла. Ее основная идея состоит в том, что синапс между двумя нейронами A и C становится тем более эффективным, чем чаще за возбуждением нейрона A следует возбуждение нейрона C, независимо от того, чем это последнее будет вызвано (рис. 144). Таким образом, для повышения эффективности синапса вовсе не обязательно, чтобы разряд нейрона C был следствием разряда A. Предположим, например, что с C образует синапс еще один нейрон, B, причем синапс A–>C слаб, а синапс В–>C весьма эффективен; допустим далее, что A и B разряжаются почти одновременно или же B чуть раньше A и что C дает разряд не под влиянием A, а в результате сильного воздействия B. В модели Хебба уже сам факт, что C разряжается сразу после A, повышает эффективность синапса А–>С. Мы предположим также, что если импульсы, приходящие к C от A, не сопровождаются импульсами в C, то синапс A–>C ослабевает.
 Рис. 144. Клетка C получает входы от клетки левого глаза A и от клетки правого глаза B. Согласно модели синапса Хебба, если разряды клетки C следуют за разрядами клетки A, то такая последовательность событий стремится усилить синапс A–>C.
Рис. 144. Клетка C получает входы от клетки левого глаза A и от клетки правого глаза B. Согласно модели синапса Хебба, если разряды клетки C следуют за разрядами клетки A, то такая последовательность событий стремится усилить синапс A–>C.
Применим эту модель к бинокулярной конвергенции у нормального животного. Пусть клетка C — бинокулярная, аксон A идет от не доминантного, а аксон B — от доминантного глаза. Недоминантный глаз будет реже активировать клетку C, чем доминантный. Согласно гипотезе Хебба, синапс A–>C будет сохраняться или усиливаться, пока за импульсами в A будут следовать импульсы в C, а такая последовательность более вероятна, если в надлежащий момент времени от другого глаза по аксону B постоянно поступает «подкрепление». А именно это и будет происходить при точном совмещении изображений на обеих сетчатках. Если активность в A не сопровождается активностью в C, синапс A–>C за длительный период времени ослабеет. Получить прямое доказательство, что модель Хебба применима к косоглазию, — дело нелегкое (по крайней мере в ближайшем будущем), но сама идея, мне кажется, заслуживает внимания.
Анатомические последствия депривации
То, что в коленчатых телах, где нет или почти нет возможностей для конкуренции глаз, мы не смогли обнаружить заметных физиологических дефектов, казалось, подтверждает идею о том, что последствия закрытия одного глаза связаны с конкуренцией, а не с бездействием глаза. Правда, клетки коленчатого тела были атрофичными, но — рассуждали мы — вряд ли можно ожидать, что все будет ясно. Если конкуренция и в самом деле играет важную роль, то корковый слой 4C мог, как нам казалось, быть подходящим объектом для проверки этой идеи, поскольку клетки в нем тоже монокулярны и конкуренция маловероятна, так что чередование полосок, связанных с левым и с правым глазом, должно остаться ненарушенным. Итак, путем длинных микроэлектродных проходок через слой 4C мы начали выяснять, сохраняются ли такие полоски после закрытия одного глаза и остаются ли нормальными их размеры. Вскоре стало ясно, что слой 4C по-прежнему подразделяется на участки левого и правого глаз, как у нормальных животных, и что клетки в полосках, соединенных с ранее закрытым глазом, остались практически нормальными. Однако последовательности клеток с доминированием закрытого глаза оказались очень короткими, как если бы полоски были аномально узкими (около 0,2 мм вместо 0,4 или 0,5 мм), а полоски, принадлежащие открытому глазу, — соответственно более широкими. Как только это стало доступным, мы начали использовать методику с инъекцией метки в глаз и транснейронным переносом ее в кору, чтобы получить прямое и наглядное подтверждение сделанных выводов. По прошествии периода депривации длительностью в несколько месяцев мы вводили кошке или обезьяне в тот или в другой глаз радиоактивную аминокислоту. Радиоавтографы показали заметное сужение полосок, связанных с ранее депривированным глазом, и соответственное расширение полосок, принадлежащих нормальному глазу. Микрофотография слева на рис. 145 иллюстрирует результаты инъекции радиоактивной аминокислоты в нормальный глаз. На этом снимке, сделанном, как обычно, в условиях темного поля, представлен срез, параллельный поверхности коры и проходящий через слой 4C. Узкие, прерывающиеся черные полоски соответствуют глазу, который был закрыт, а более широкие светлые (меченые) полоски — открытому глазу, в который ввели метку. На рис. 146 показана обратная картина, полученная после введения метки в ранее закрытый глаз. Этот срез был сделан поперек слоя 4C, поэтому мы видим «полоски» в поперечном сечении.

 Рис. 145. Срезы зрительной коры макака, правый глаз которого был закрыт с момента рождения до 18-месячного возраста. В левый глаз инъецировали радиоактивную аминокислоту, и неделю спустя приготовили срезы мозга, параллельные поверхности зрительной коры. Кора имеет куполообразную форму, поэтому такие срезы вначале идут тангенциально, а затем образуют кольца (А) все большего диаметра, подобные кольцам луковицы. Эти кольца были вырезаны из микрофотографий и склеены вместе (Б). Впоследствии мы научились расправлять кору перед ее замораживанием, и тогда уже не приходилось разрезать и склеивать фотографии серийных срезов. На обычном радиоавтографе зерна серебра выглядят черными на белом фоне. Мы, однако, применяли метод исследования в темном поле, при котором зерна серебра рассеивают свет и их скопления выглядят как светлые участки. Светлые полоски — места поступления в слой 4C меченых атомов из открытого глаза — расширены, а темные (связанные с закрытым глазом) сильно сужены.
Рис. 145. Срезы зрительной коры макака, правый глаз которого был закрыт с момента рождения до 18-месячного возраста. В левый глаз инъецировали радиоактивную аминокислоту, и неделю спустя приготовили срезы мозга, параллельные поверхности зрительной коры. Кора имеет куполообразную форму, поэтому такие срезы вначале идут тангенциально, а затем образуют кольца (А) все большего диаметра, подобные кольцам луковицы. Эти кольца были вырезаны из микрофотографий и склеены вместе (Б). Впоследствии мы научились расправлять кору перед ее замораживанием, и тогда уже не приходилось разрезать и склеивать фотографии серийных срезов. На обычном радиоавтографе зерна серебра выглядят черными на белом фоне. Мы, однако, применяли метод исследования в темном поле, при котором зерна серебра рассеивают свет и их скопления выглядят как светлые участки. Светлые полоски — места поступления в слой 4C меченых атомов из открытого глаза — расширены, а темные (связанные с закрытым глазом) сильно сужены.
 Рис. 146. Другой обезьяне метку ввели в закрытый глаз. Срез сделан в поперечном, а не в тангенциальном направлении. На этом снимке, полученном в темном поле, светлые полоски в слое 4C (которые видны здесь в поперечном сечении) сильно сужены.
Рис. 146. Другой обезьяне метку ввели в закрытый глаз. Срез сделан в поперечном, а не в тангенциальном направлении. На этом снимке, полученном в темном поле, светлые полоски в слое 4C (которые видны здесь в поперечном сечении) сильно сужены.
Результаты, полученные на слое 4C, подкрепили наши сомнения относительно конкурентной модели, связанные с фактом атрофии клеток коленчатого тела: либо гипотеза о конкуренции была неверна, либо какая-то погрешность была в наших рассуждениях. Оказалось, что погрешность касалась как коленчатого тела, так и коры. В коре нашей ошибкой было предположение, что к моменту, когда мы закрывали глаза новорожденным животным, колонки глазодоминантности были уже вполне сформированы.
Нормальное развитие колонок глазодоминантности
Очевидный способ получения сведений о колонках глазодоминантности у новорожденных животных заключался в проверке распределения волокон, входящих в слой 4C, путем инъекции метки в глаз на первый или второй день жизни. Результат оказался неожиданным. Вместо четких, ясно очерченных полосок в слое 4C мы наблюдали непрерывное распределение метки. Радиоавтограф слева на рис. 147 показывает этот слой в поперечном разрезе, и мы не видим никаких признаков колонок. Только если мы нареза́ли кору параллельно ее поверхности, можно было увидеть слабую волнистость с полумиллиметровыми интервалами, как это показано на радиоавтографе справа. Очевидно, волокна, растущие от коленчатого тела в кору, не разветвляются сразу же к областям, связанным с левым и с правым глазом. Они сначала посылают свои ветви повсюду в радиусе нескольких миллиметров и лишь позднее, приблизительно к моменту рождения животного, втягиваются обратно и окончательно распределяются. Слабая волнистость у новорожденного показывает, что втягивание началось еще до рождения; Пашко Ракич с большими трудностями сумел инъецировать метку в глаз утробного плода обезьяны и выяснил, что оно начинается за несколько недель до рождения. Инъецируя метку в один глаз в различные моменты после рождения, мы смогли легко установить, что в первые две или три недели в слое 4 происходит постепенное втягивание окончаний, так что к четвертой неделе формирование полосок завершается. Узор полосок и их периодичность с шагом 0,8 миллиметра, таким образом, являются врожденными.
 Рис. 147. Слева: поперечный срез слоя 4C коры новорожденного макака после инъекции радиоактивной метки в глаз. Снимок сделан в темном поле, поэтому меченые участки выглядят светлыми. Их непрерывность показывает, что входы от каждого глаза не сгруппированы в полоски, а перемешаны во всем слое. (Белая полоса между слоями 4C, находящимися у поверхности мозга и в глубине, представляет собой белое вещество мозга, заполненное волокнами с меткой, идущими от наружных коленчатых тел.)
Справа: здесь показан срез другого полушария; при изготовлении среза нож слегка коснулся участка стриарной коры, находящегося в глубине. В верхней части слоя 4C можно усмотреть намек на полоски. (Эти полоски находятся в подразделении, связанном с крупноклеточными слоями коленчатого тела.) Более глубинный подслой β образует непрерывное кольцо вокруг α и поэтому предположительно позднее дифференцируется по глазодоминантности.
Рис. 147. Слева: поперечный срез слоя 4C коры новорожденного макака после инъекции радиоактивной метки в глаз. Снимок сделан в темном поле, поэтому меченые участки выглядят светлыми. Их непрерывность показывает, что входы от каждого глаза не сгруппированы в полоски, а перемешаны во всем слое. (Белая полоса между слоями 4C, находящимися у поверхности мозга и в глубине, представляет собой белое вещество мозга, заполненное волокнами с меткой, идущими от наружных коленчатых тел.)
Справа: здесь показан срез другого полушария; при изготовлении среза нож слегка коснулся участка стриарной коры, находящегося в глубине. В верхней части слоя 4C можно усмотреть намек на полоски. (Эти полоски находятся в подразделении, связанном с крупноклеточными слоями коленчатого тела.) Более глубинный подслой β образует непрерывное кольцо вокруг α и поэтому предположительно позднее дифференцируется по глазодоминантности.
Мы легко подтвердили представление о постнатальном втягивании окончаний, регистрируя ответы слоя 4C обезьян вскоре после рождения. По мере продвижения электрода вдоль этого слоя параллельно его поверхности мы везде смогли вызвать активность от обоих глаз вместо четкого чередования глаз, наблюдаемого у взрослых животных. Карла Шатц продемонстрировала аналогичный процесс развития коленчатого тела у кошки: у утробного плода многие клетки коленчатого тела временно получают входы от обоих глаз, но утрачивают один из входов по мере образования слоистой структуры. Таким образом, и в коре, и в коленчатом теле мы имеем примеры формирования и последующей спонтанной дегенерации синапсов в процессе развития. Окончательная картина чередования полос для левого и правого глаза в корковом слое 4C развивается нормально даже в том случае, если оба глаза закрыты путем сшивания век; это означает, что надлежащие связи могут создаваться и без участия индивидуального опыта. Мы полагаем, что в процессе развития входные волокна от обоих глаз конкурируют в слое 4C таким образом, что если входы от одного из глаз в каком-то месте преобладают, то преимущество этого глаза имеет тенденцию увеличиваться, а число входов от другого глаза соответственно убывает. Даже небольшое начальное неравенство при этом стремится постепенно возрастать до тех пор, пока всюду в слое 4 в возрасте одного месяца не появятся отчетливые полоски с полным доминированием. При закрытом глазе баланс нарушается, и на границах полосок, где в норме исход борьбы решает прямое столкновение, открытый глаз получает преимущество и одерживает верх, что показано на рис. 148. Мы не знаем, что́ при нормальном развитии приводит к начальному дисбалансу, но при столь неустойчивом равновесии дело может решить даже мельчайшее различие. Почему развивающийся узор должен состоять из параллельных полосок шириной 0,5 мм каждая, остается предметом догадок. Некоторые исследователи предполагают, что аксоны от одного и того же глаза, сближаясь, начинают притягивать друг друга, а от разных глаз — отталкивать друг друга, причем на малых расстояниях отталкивание слабее притяжения, так что притяжение побеждает. С увеличением расстояния притягивающая сила убывает быстрее отталкивающей, так что на некотором удалении побеждает отталкивание. Шириной зоны этих конкурентных взаимодействий и определяются размеры колонок. Согласно математическим расчетам, для получения параллельных полосок (в противовес шахматному порядку или островкам аксонов от левого глаза в сплошной массе аксонов от правого глаза) требуется лишь то, чтобы границы между колонками были как можно короче. Итак, мы имеем возможность объяснить сужение и расширение колонок, показав, что в то время, когда глаз был закрыт в раннем периоде жизни, конкуренция все же была возможной.
 Рис. 148. Эта «конкурентная модель» объясняет разделение волокон четвертого слоя на колонки глазодоминантности. К моменту рождения колонки уже начали формироваться. В норме, если в какой-то точке один глаз хотя бы незначительно доминирует, это оканчивается его полной монополией. Если один глаз при рождении закрыт, то сохранившиеся волокна от открытого глаза в любой данной точке слоя 4 полностью одерживают верх. Волокна от закрытого глаза сохраняются только там, где в момент закрытия у них не было конкурентов.
Рис. 148. Эта «конкурентная модель» объясняет разделение волокон четвертого слоя на колонки глазодоминантности. К моменту рождения колонки уже начали формироваться. В норме, если в какой-то точке один глаз хотя бы незначительно доминирует, это оканчивается его полной монополией. Если один глаз при рождении закрыт, то сохранившиеся волокна от открытого глаза в любой данной точке слоя 4 полностью одерживают верх. Волокна от закрытого глаза сохраняются только там, где в момент закрытия у них не было конкурентов.
Тем временем Рэй Гиллери, работавший тогда в Висконсинском университете, предложил правдоподобное объяснение атрофии клеток в коленчатом теле. Изучая наши рисунки, демонстрирующие сморщивание клеток после закрытия одного глаза у кошек, он заметил, что в участках коленчатого тела, наиболее удаленных от средней линии, сморщивание было значительно меньшим; действительно, там — в проекции височной серповидной области — клетки казались вполне нормальными. Здесь отображается боковой край зрительного поля, настолько удаленный от середины, что его может видеть лишь один глаз этой стороны (рис. 149). Мы были огорчены, если не сказать больше, ибо настолько увлеклись измерением клеток с целью подтвердить наши выводы, что просто забыли как следует рассмотреть собственные микрофотографии. Ведь отсутствие атрофии клеток со входами от «височного серпа» означало, что атрофия в других местах коленчатого тела, и в самом деле, могла быть результатом конкуренции со стороны другого глаза.
 Рис. 149. Различные участки обеих сетчаток проецируются на их собственные зоны в правом коленчатом теле кошки (которое показано здесь в поперечном сечении). Верхний слой коленчатого тела, получающий входные волокна от контралатерального (левого) глаза, нависает над следующим слоем. Свешивающаяся часть получает входы от височного серповидного участка поля зрения, который проецируется на сетчатку только одного глаза. При закрытии этого глаза свешивающаяся часть не атрофируется — предположительно потому, что не встречает конкуренции со стороны другого глаза, в котором соответственного участка нет.
Рис. 149. Различные участки обеих сетчаток проецируются на их собственные зоны в правом коленчатом теле кошки (которое показано здесь в поперечном сечении). Верхний слой коленчатого тела, получающий входные волокна от контралатерального (левого) глаза, нависает над следующим слоем. Свешивающаяся часть получает входы от височного серповидного участка поля зрения, который проецируется на сетчатку только одного глаза. При закрытии этого глаза свешивающаяся часть не атрофируется — предположительно потому, что не встречает конкуренции со стороны другого глаза, в котором соответственного участка нет.
С помощью весьма остроумного эксперимента, показанного на рис. 150, Маррей Шерман и его коллеги представили решающие доказательства роли конкуренции в атрофии клеток коленчатого тела. Сначала они разрушили у котенка крошечный участок одной сетчатки в области бинокулярного поля зрения. Затем они плотно зашили другой глаз. В небольшой зоне того слоя коленчатого тела, на который проецировался глаз с локальным повреждением, была обнаружена сильно выраженная атрофия. Этот результат получили и многие другие исследователи. В слое, имевшем входы от другого, ранее закрытого глаза, как и ожидалось, они тоже наблюдали сморщивание клеток повсюду, кроме зоны, подвергшейся атрофии в слое с проекцией другого глаза. Там, несмотря на отсутствие входных сигналов от глаза, клетки были нормальными. Атрофия, обусловленная закрытием глаза, была предотвращена путем исключения конкуренции. Ясно, что конкуренция не могла происходить в самом коленчатом теле; однако следует помнить, что, хотя тела и дендриты клеток коленчатого тела находились в самом коленчатом теле, большинство окончаний их аксонов были в коре, а там, как я уже говорил, окончания, принадлежащие закрытому глазу, подвергались сильной атрофии. Вывод состоит в том, что при закрытии глаза сморщивание клеток — это результат уменьшения числа поддерживаемых ими аксонных окончаний.
 Рис. 150. В 1974 году эксперимент Шермана, Гиллери, Кааса и Сандерсона продемонстрировал значение конкуренции для атрофии клеток наружного коленчатого тела. Если разрушить у котенка небольшой участок левой сетчатки, то в соответствующей части верхнего слоя правого наружного коленчатого тела образуется островок сильно выраженной атрофии. Если затем закрыть правый глаз, то, как и можно было ожидать, слой, расположенный ниже, подвергается атрофии, за исключением участка, лежащего прямо под атрофированным участком верхнего слоя. Этот факт служит сильным доводом в пользу конкурентной природы атрофии, вызываемой закрытием глаза.
Рис. 150. В 1974 году эксперимент Шермана, Гиллери, Кааса и Сандерсона продемонстрировал значение конкуренции для атрофии клеток наружного коленчатого тела. Если разрушить у котенка небольшой участок левой сетчатки, то в соответствующей части верхнего слоя правого наружного коленчатого тела образуется островок сильно выраженной атрофии. Если затем закрыть правый глаз, то, как и можно было ожидать, слой, расположенный ниже, подвергается атрофии, за исключением участка, лежащего прямо под атрофированным участком верхнего слоя. Этот факт служит сильным доводом в пользу конкурентной природы атрофии, вызываемой закрытием глаза.
Открытие того факта, что при рождении во всем слое 4 без каких-либо перерывов представлены волокна от обоих глаз, оказалось весьма полезным, так как оно объясняло, каким образом могла бы реализоваться конкуренция на синаптическом уровне в структуре, в которой, казалось бы, нет никаких возможностей для взаимодействия глаз. И все-таки проблема может оказаться не такой простой. Если причина изменений в слое 4 — то, что в первые недели после рождения здесь существуют условия для конкуренции, то закрытие глаза в возрасте, когда система еще пластична, а колонки уже разделены, не приводило бы к возникновению изменений. Мы закрывали глаз в возрасте пяти с половиной недель и только через год вводили метку в другой глаз. Результатом было четкое сужение и расширение соответствующих полос. Это как будто бы указывает на то, что помимо дифференцированного втягивания окончаний возможно и прорастание их на новую территорию.
Другие специальные эксперименты с депривацией
Во всех описанных до сих пор исследованиях мы закрывали один или оба глаза или же перереза́ли наружные мышцы одного из глаз. Вскоре во многих лабораториях было проведено множество других экспериментов, включающих чуть ли не все мыслимые виды зрительной депривации. В одном из первых и наиболее интересных экспериментов ставился вопрос, будет ли содержание животного в условиях, позволяющих ему видеть полоски лишь одной ориентации, приводить к утрате клеток, чувствительных ко всем иным ориентациям. В 1970 году Колин Блейкмор и Дж. Ф. Купер из Кембриджского университета ежедневно с раннего возраста показывали котятам в течение нескольких часов чередующиеся черные и белые вертикальные полосы, а в остальное время содержали их в темноте. Результатом было сохранение корковых клеток, реагирующих на вертикальные полосы, и резкое уменьшение числа клеток, предпочитающих другие ориентации. Неясно, перестали ли клетки с исходно промежуточными ориентациями отвечать вовсе или они сменили предпочитаемое направление на вертикальное. В опубликованной в том же году работе Хельмут Хирш и Нико Спинелли использовали очки, позволявшие котенку видеть одним глазом только вертикальные, а другим — только горизонтальные контуры. В результате получилась кора, содержащая клетки с предпочтением вертикалей, клетки с предпочтением горизонталей, но очень мало клеток, предпочитающих наклонные линии. Кроме того, на клетки, активируемые горизонтальными линиями, влиял лишь тот глаз, который подвергался раньше воздействию горизонтальных линий, а на клетки, возбуждаемые вертикальными линиями, — лишь глаз, подвергавшийся воздействию вертикальных линий. Другие интересные процедуры включали выращивание животных в темном помещении, в котором один или несколько раз в секунду вспыхивал яркий импульс света; он позволял животному увидеть, где оно находится, но должен был сводить к минимуму восприятие любого движения. Результатом этих экспериментов, которые проводили в 1975 году Макс Цинадер, Нэнси Берман и Алан Хейн в Массачусетсском технологическом институте, а также М. Цинадер и Г. Черненко в Далхауси (Галифакс), было уменьшение числа клеток, чувствительных к движению. В другой серии экспериментов, которую начали Ф. Треттер, М. Цинадер и Вольф Зингер в Мюнхене, животным показывали только движение полосок слева направо и получили ожидаемое асимметричное распределение в коре клеток, чувствительных к направлению движения. С большими трудностями и затратами мы с Торстеном Визелом выращивали детеныша обезьяны в комнате, освещаемой лишь длинноволновым красным светом, а затем регистрировали ответы клеток наружных коленчатых тел, чтобы выяснить, не окажется ли здесь необычно мало цветокодирующих нейронов (см. гл. 8). Какой-либо аномалии в коленчатых телах обнаружить не удалось. Наконец, в последнее десятилетие появилось множество исследований, имеющих целью узнать, содержат ли модифицируемые синапсы такие специальные нейромедиаторы или нейромодуляторы, как норадреналин, а также ацетилхолин или серотонин. Результаты этих исследований представят большой интерес.
Значение результатов депривации в более широкой перспективе
Часто спрашивают, какой цели служит пластичность зрительной системы в ранний период жизни (у человека период чувствительности предположительно составляет четыре — пять лет). У животных, у которых один глаз зашит с момента рождения, территория открытого глаза в слое 4C расширяется; дает ли это какое-либо преимущество открытому глазу? Ответа на этот вопрос еще нет. Трудно представить себе, что острота зрения становится лучше, чем в норме, поскольку нормальная острота, измеряемая офтальмологом с помощью испытательной таблицы, определяется в конечном счете плотностью упаковки рецепторов (см. гл. 3), а она уже лимитирована длиной световых волн. Как бы то ни было, кажется весьма маловероятным, чтобы пластичность выработалась в ходе эволюции всего лишь на случай возможной утраты глаза детенышем или появления у него косоглазия. Популярная и, несомненно, правдоподобная идея состоит в том, что пластичность обеспечивает тонкую настройку связей, необходимую для восприятия форм, движения и глубины, и что эта настройка осуществляется в основном постнатально под контролем самого зрения. Эта идея привлекательна потому, что способность мозга обучаться может избавить его от необходимости заранее программировать все детали и может обеспечить достаточную гибкость для адаптации к разнообразным условиям среды. Однако для такой идеи пока еще нет убедительных экспериментальных подтверждений. Я лично склонен думать, что в первичной зрительной коре и, быть может, на нескольких последующих уровнях связи полностью определяются генетическими инструкциями. Ясно, что значительная часть этих связей образуется пренатально и, следовательно, без участия сенсорного опыта, и какая бы стратегия ни использовалась для их построения, она могла бы также обеспечивать и их тонкую настройку. Я не утверждаю, что так же обстоит дело и в других областях коры. Большинство неврологов полагают, что нейронные цепи, ответственные за овладение языком, находятся в основном в коре, — и никто не станет утверждать, что мы рождаемся со знанием тонкостей нашего родного языка. Модифицируемость различных корковых зон и возрастной диапазон, в пределах которого возможны модификации, могут широко варьировать от зоны к зоне, причем первичная зрительная кора с ее минимальной пластичностью, ограниченной наименьшим числом лет постнатального развития, видимо, являет собой крайний случай. На более периферических уровнях зрительной системы, включающих сетчатку, коленчатое тело и первичную кору, пластичность может быть просто побочным продуктом созревания; здесь, я полагаю, определенную роль играет конкуренция, и помехи в ее осуществлении ведут к искажению связей. Но в действительности мы еще не знаем, почему зрительная кора может быть модифицирована в раннем периоде жизни. Стоит отметить, что все модификации в наших экспериментах явились результатом аномального раннего опыта и привели к образованию аномальных связей. Эксперименты, имевшие целью показать, что обогащенный ранний опыт ведет к усиленному развитию коры или иных структур мозга, не кажутся мне очень убедительными. Что больше всего поразило нас в экспериментах со зрительной депривацией, так это возможность вызывать заметные физиологические и морфологические изменения в нервной системе без реального физического вмешательства. Давно известно, что клетки в нервной системе могут дегенерировать, если нерв физически перерезать или раздавить, но в описанных мною экспериментах все воздействие сводилось к исключению света, а в опытах с косоглазием вмешательство было еще более тонким. В каждом случае «наказание» более или менее соответствует «преступлению». Исключите форму, и клетки, в норме отвечающие только на формы, перестанут на них отвечать. Измените относительное положение глаз, перерезав одну из мышц, и связи, в норме обслуживающие бинокулярные взаимодействия, будут разорваны. Устраните подвижные стимулы вообще или движение их в определенном направлении, и соответствующие клетки не будут реагировать. Вряд ли нужно обладать каким-то особым воображением, чтобы подумать о том, что у ребенка, лишенного социальных контактов, или у выращенного в изоляции животного, как в некоторых экспериментах Гарри Харлоу, могут возникнуть аналогичные, столь же реальные изменения в зонах мозга, определяющих эмоции или взаимодействия с другими особями того же вида. Конечно, ни один патолог изменений пока не наблюдал, но ведь и после зрительной депривации в коре тоже нельзя без весьма специальных методов (вроде инъекции метки и глаз) увидеть какие-либо изменения. Когда одни аксоны укорачиваются, а другие удлиняются, вся структура даже при исследовании в электронном микроскопе выглядит точно так же. Таким образом, возможное значение экспериментов с депривацией выходит далеко за рамки зрительной системы — оно касается также неврологии в целом и в значительной мере психиатрии. Фрейд вполне мог быть прав, связывая психоневрозы с событиями раннего детства, и, учитывая его неврологическое образование, я думаю, он был бы в восторге от мысли, что подобные события могут приводить к заметным гистологическим или гистохимическим изменениям в реальном физическом мозгу.
10. Настоящее и будущее
При создании этой книги я намеревался описать то, что нам известно об анатомии и физиологии путей зрительной системы вплоть до стриарной коры. Сведения, которыми мы ныне располагаем, — это, по-существу, лишь первые плоды наших попыток понять физиологическую основу восприятия, лишь начало увлекательной повести, следующие главы которой еще только-только просматриваются; со средней дистанции мы можем видеть лишь главные горные хребты, конец их — за пределами видимости. Стриарная кора — всего только первая из более чем дюжины отдельных зрительных зон, в каждой из которых представлено все поле зрения. Вместе эти зоны образуют подобие лоскутного одеяла, которое покрывает затылочную кору и простирается вперед на заднюю височную и заднюю теменную кору. Начиная со стриарной коры, каждая зона снабжает информацией две или несколько вышележащих (в смысле иерархии) зон, а связи между ними топографически организованы так, что любая данная зона, подобно стриарной коре, содержит упорядоченное отображение поля зрения. Восходящие связи предположительно передают зрительную информацию из одной области в другую для дальнейшей обработки. Наша задача состоит в том, чтобы для каждой из этих зон выяснить, каким образом здесь обрабатывается информация, — та же задача, с которой мы сталкивались ранее, интересуясь, что делает стриарная кора с информацией, получаемой от коленчатого тела. Хотя мы лишь недавно узнали о том, как многочисленны эти зрительные зоны, уже накапливаются данные о связях и физиологии одиночных клеток некоторых из них. Подобно тому как поле 17 является мозаикой двух совокупностей областей, пузырьковой и непузырьковой, следующая зрительная зона — поле 18, или зрительная зона 2, — состоит из трех совокупностей. В отличие от пузырьков и непузырьков, образующих как бы островки в океане, мозаика поля 18 имеет форму параллельных полосок. В системе этих полосок мы находим удивительное разделение функций. В совокупности толстых полосок большинство клеток весьма чувствительно к относительному горизонтальному положению стимулов, подаваемых на оба глаза, как это описано в главе 7; отсюда мы заключаем, что это подразделение с толстыми полосками связано, по крайней мере отчасти, со стереоскопическим зрением. У клеток второй совокупности, расположенных в тонких полосках, отсутствует ориентационная избирательность, и они часто демонстрируют специфические цветовые реакции. Клетки третьей совокупности, расположенные в бледных полосках, ориентационно-избирательны и по большей части реагируют на концы линий. Таким образом, три совокупности подразделений, составляющие поле 18, связаны, видимо, со стереопсисом, цветом и формой. Аналогичное разделение функций имеется и в областях, расположенных выше поля 18, но здесь каждая зона в целом, по-видимому, выполняет лишь одну или, быть может, две зрительные функции. Зона, называемая MT (от middle temporal gyrus — средняя височная извилина), связана с восприятием движения и стереопсисом, а зона V4 — вероятно, в основном с цветом. Таким образом, мы можем различить два идущих рука об руку процесса. Первый из них носит иерархический характер. Для решения различных зрительных задач, обрисованных в предыдущих главах и относящихся к цвету, стереопсису, движению и форме, в обработку информации включается одна зона за другой, и в этом ряду все больше возрастают степень абстрагирования и сложность отображения. Второй процесс состоит в дивергенции путей. По-видимому, решаемые задачи требуют столь различных стратегий и нервных механизмов, что становится выгоднее распределять их между обособленными каналами. Рис. 151. Отбивающий в бейсболе поворачивается к мячу, движущемуся к нему со скоростью около 100 миль в час; при такой большой скорости весь полет мяча длится около секунды. Результат действий игрока определяется несколькими миллиметрами рассогласования. Успех или неудача зависит от работы зрительных структур — тех самых, которые рассматривались в этой книге, и многочисленных других, расположенных на более высоких уровнях зрительной системы, а также от двигательных структур, включающих моторную кору, мозжечок, мозговой ствол и спинной мозг.
Рис. 151. Отбивающий в бейсболе поворачивается к мячу, движущемуся к нему со скоростью около 100 миль в час; при такой большой скорости весь полет мяча длится около секунды. Результат действий игрока определяется несколькими миллиметрами рассогласования. Успех или неудача зависит от работы зрительных структур — тех самых, которые рассматривались в этой книге, и многочисленных других, расположенных на более высоких уровнях зрительной системы, а также от двигательных структур, включающих моторную кору, мозжечок, мозговой ствол и спинной мозг.
Эта удивительная тенденция обрабатывать такие признаки, как форма, цвет и движение, в отдельных структурах мозга сразу же вызывает вопрос: каким образом вся эта информация в конце концов собирается вместе при восприятии, скажем, прыгающего красного мячика? Ясно, что она должна собраться где-то воедино — хотя бы только в двигательных нервах, управляющих актом схватывания. Где она собирается и как это происходит, мы не имеем ни малейшего представления.
Вот здесь мы и находимся сейчас, в 1987 году, в нашем пошаговом анализе зрительного пути. По числу синапсов (около 8 или 10) и сложности преобразований путь от палочек и колбочек сетчатки до зон MT или зрительной зоны 2 в коре может показаться длинным, но, несомненно, еще намного длиннее путь от таких процессов, как ответ на ориентацию, концы линий, диспаратность или соотношение оппонентных цветов, к распознаванию любых форм, воспринимаемых нами в повседневной жизни. Мы далеки от того, чтобы понимать восприятие объектов, даже таких сравнительно простых, как круг, треугольник или буква A, — мы далеки даже от возможности высказывать правдоподобные гипотезы на этот счет. Нам не следует испытывать особого удивления или смущения по поводу нашего невежества в отношении этих тайн. Работающие в области искусственного интеллекта (ИИ) не могут создать машину, которая начала бы соперничать с мозгом в выполнении таких специальных задач, как чтение рукописного текста, вождение автомобиля или узнавание лиц. Они, однако, показали, что решение любой из этих задач связано с огромными теоретическими трудностями. Дело не в непреодолимости этих трудностей — ведь мозг с ними явно справляется; речь идет скорее о том, что применяемые мозгом методы не могут быть простыми: на языке ИИ говорят, что задачи «нетривиальны». Итак, мозг решает нетривиальные задачи. Удивительно то, что он решает не две или три, а тысячи таких задач. После лекции, когда наступает время отвечать на вопросы, специалиста по сенсорной физиологии или психологии часто спрашивают, каковы наиболее правдоподобные предположения относительно того, каким образом распознаются видимые объекты. Становятся ли клетки при переходе к более центральным уровням все более и более специализированными, так что на каком-то уровне могут найтись клетки, реагирующие на лицо одного-единственного конкретного человека — например, чьей-то бабушки? Такое представление, называемое теорией бабушкиной клетки, вряд ли можно принимать всерьез. Можем ли мы обнаружить отдельные клетки для бабушки улыбающейся, плачущей или занимающейся шитьем? Или отдельные клетки, отражающие понятие или определение «бабушки» — то, что это мать матери или мать отца какого-то человека? И если бы у нас действительно имелись «бабушкины клетки», что тогда? Куда они посылали бы свои выходные сигналы? Альтернативой служит предположение, что данный объект активирует определенную группу клеток — «нейронный ансамбль», каждый член которого может принадлежать также и другим ансамблям. Поскольку известно, что разрушение небольшого участка мозга обычно не ведет к исчезновению определенных воспоминаний, приходится предполагать, что клетки одного ансамбля не сосредоточены в одной корковой зоне, а разбросаны по многим зонам. Таким образом, «бабушке,занимающейся шитьем», должен соответствовать более крупный ансамбль, включающий бабушку по определению, бабушкино лицо и процесс шитья. Понятно, что придумать способ экспериментальной проверки подобных представлений непросто. Даже в стриарной коре нелегко зарегистрировать ответы одиночной клетки и осмыслить результаты; тем более трудно вообразить, каким образом можно постичь работу клетки, входящей, быть может, в сотню ансамблей, каждый из которых содержит тысячу клеток. Попытавшись однажды регистрировать активность трех клеток одновременно и понять их совместную роль в повседневной жизни животного, я могу лишь восхищаться усилиями тех, кто надеется создать системы электродов для одновременной регистрации импульсов в сотнях клеток. Но теперь мы привыкаем уже иметь дело с задачами, трудность которых еще вчера казалась непреодолимой. Текущий счет в пользу смутных представлений о клеточных ансамблях открыт уже давно, и на него продолжают поступать данные о наличии корковых областей, специализированных для восприятия лиц. Группа Чарлза Гросса из Принстона обнаружила в мозгу обезьяны в одной из зрительных зон височной доли клетки, как будто бы избирательно реагирующие на лица. Люди с повреждениями определенного участка в нижней области затылочной доли нередко утрачивают способность узнавать лица, в том числе даже лица близких родственников. Антонио Дамасио из Университета Айовы высказал предположение, что эти больные неспособны узнавать не только лица, но и более широкий класс объектов, включающий лица. Он описывает женщину, которая помимо лиц не узнавала также индивидуальные автомашины. Она могла отличить легковой автомобиль от грузовика, но, разыскивая собственную машину на стоянке, она была вынуждена обходить ряды машин и читать их номерные знаки, что указывает на интактность как ее зрения, так и способности читать номера. Построение умозрительных гипотез может быть делом весьма увлекательным, но когда все-таки можем мы надеяться получить ответы на некоторые вопросы, связанные с восприятием? Больше 35 лет прошло с тех пор, когда Куффлер изучил свойства ганглиозных клеток сетчатки. За это время наши представления о сложности зрительных путей и о круге проблем, касающихся восприятия, радикально изменились. Мы поняли, что открытие рецептивных полей с центром и периферией и клеток с ориентационной избирательностью — это лишь два этапа в разгадке головоломки, содержащей сотни подобных этапов. Мозг, если даже говорить только о зрении, предназначен для выполнения многих задач, и миллионы лет эволюции привели к выработке весьма остроумных решений. В результате упорной работы мы можем прийти к пониманию любой небольшой их части, но вряд ли мы сможем разобраться во всем. Было бы так же нереалистично верить, что когда-нибудь мы сможем понять сложную работу каждого из миллионов белков, существующих в нашем организме. С философских позиций, однако, важно располагать хотя бы несколькими примерами (идет ли речь о нейронных структурах или белках), которые бы мы хорошо понимали: наша способность разгадать даже малую часть процессов, связанных с жизнью — или с восприятием, мышлением, эмоциями, — говорит нам, что полное понимание принципиально возможно и что мы не нуждаемся в апелляции к мистическим жизненным силам или к душе. У некоторых людей может возникнуть опасение, что такого рода материалистическая концепция, рассматривающая мозг как некую супермашину, лишит нашу жизнь очарования и отвратит нас от духовных ценностей. Это сродни опасению, что знание анатомии человека помешает нам восхищаться формами человеческого тела. Художники и медики знают, что верно как раз обратное. Проблема здесь лингвистическая: если машиной называют нечто с заклепками, храповиками и шестернями, то это действительно звучит неромантично. Но я разумею под машиной всякий объект, выполняющий свои функции в согласии с законами физики, объект, который мы можем в конечном счете понять точно таким же образом, как мы понимаем работу печатного станка. Я верю, что мозг — именно такой объект. Надо ли тревожиться о возможных ужасных последствиях понимания механизмов мозга, аналогичных последствиям понимания атома? Нужно ли опасаться того, что ЦРУ станет читать или контролировать наши мысли? Я не вижу оснований, чтобы мучиться по этому поводу бессонницей, по крайней мере в предстоящем столетии. Из всех предшествующих глав этой книги должно быть очевидно, что чтение мыслей или управление ими с помощью достижений нейрофизиологии примерно столь же осуществимо, как поездка на уик-энд в галактику Андромеды и обратно. Но даже если бы контроль над мыслями оказался в принципе возможным, по сравнению с ним профилактика или излечение миллионов страдающих шизофренией должны быть легкой задачей. Я предпочел бы рискнуть и продолжать исследования. Мы можем вскоре столкнуться с проблемой иного рода: как примирить некоторые из наших наиболее сокровенных и глубоко укоренившихся верований с новыми сведениями о мозге. В 1983 году римская католическая церковь официально указала на приемлемость физики и космологии, провозглашенной Галилеем 350 лет тому назад. Сегодня наши судьи, политические деятели и издатели столкнулись с аналогичной проблемой в связи с изучением школьниками фактов, касающихся эволюции и молекулярной биологии. Если дух и душа значат для нейробиологии то же, что небеса для астрономии и сотворение мира для биологии, то в недалеком будущем можно ожидать третьей революции в умах. Нам не следует, однако, самодовольно считать все это проявлением борьбы между мудростью науки и религиозным невежеством. Если люди склонны бережно хранить некоторые верования, то вполне разумно предполагать, что эволюция нашего мозга способствовала этой склонности — по причинам, связанным с выживанием. Отказ от старых верований или мифов и замена их научным мировоззрением не должны осуществляться поспешно или по указу. Но я думаю, что нам придется в конце концов видоизменить наши верования и отыскать в них место для фактов, которые наш мозг позволил нам установить путем эксперимента и дедукции: Земля круглая; она вращается вокруг Солнца; живые существа эволюционируют; жизнь можно объяснить фантастически сложным взаимодействием молекул; а мысль удастся когда-нибудь понять как функцию фантастически сложных систем нейронных соединений. Потенциальная польза от познания мозга связана не только с лечением и профилактикой неврологических и психических заболеваний. Она должна затронуть и такие, например, области, как образование, где мы тоже пытаемся влиять на мозг. Так разве мы не сможем учить лучше, если познаем объект, на который хотим воздействовать? Возможная польза распространяется даже на искусство, музыку, спорт и общественные отношения. Все, что мы делаем, зависит от нашего мозга. Сказав все это, я все-таки должен признаться, что самым сильным побудительным мотивом для меня и, полагаю, для большинства моих коллег служит чистая любознательность по отношению к работе самой сложной из известных ныне структур.
Литература для дополнительного чтения
К главе 1.
Scientific American issue on the brain, 241 (3), September 1979. Reprinted as The Brain, A Scientific American Book, W. H. Freeman, New York, 1979. Nauta W.J.H., Feirtag M. Fundamental Neuroanatomy, W. H. Freeman, New York, 1986. Ramón у Cajal, Santiago. Histologie du Système Nerveux de l’Homme et des Vertébrés, vols. 1 and 2 (translated by L. Azoulay from the Spanish), Madrid, 1952.
К главе 2.
Kandel E. R., Schwartz J. H. Principles of Neural Science, Elsevier North-Holland, New York, 1981. Kuffler S. W., Nicholls J. G., Martin A.R. From Neuron to Brain, 2d ed., Sinauer Associates, Sunderland, Mass., 1984.
К главе 3.
Dowling J. E. The Retina — An Approachable Part of the Brain, Harvard University Press, Cambridge, Mass., 1987. Kuffler S. W., Nicholls J. G., Martin A. R. From Neuron to Brain, 2d ed., Sinauer Associates, Sunderland, Mass., 1984. Kuffler S. W. Neurons in the retina: Organization, inhibition and excitatory problems. Cold Spring Harbor Symposia on Quantitative Biology, 17, 281—292 (1952). Schnapf J. L., Baylor D. A. How photoreceptor cells respond to light. Sci. Am., 256. 40—47 (1987).
К главе 4.
Hubel D. H., Wiesel T. N. Receptive fields of single neurones in the cat’s striate cortex. J. Physiol., 148. 574—591 (1959). Hubel D. H., Wiesel T. N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J. Physiol., 160. 106—154 (1962). Hubel D. H., Wiesel T. N. Receptive fields and functional architecture in two non-striate visual areas (18 and 19) of the cat. J. Neurophysiol., 28, 229—289 (1965). Hubel D. H., Wiesel T. N. Receptive fields and functional architecture of monkey striate cortex. J. Physiology, 195, 215—243 (1968). Hubel D. H., Wiesel T. N. Brain mechanisms of vision. Sci. Am., 241, 130—144 (1979). Hubel D. H. Exploration of the primary visual cortex, 1955—78 (Nobel Lecture). Nature, 299, 515—524 (1982).
К главам 5 и 6.
Hubel D. H., Wiesel T. N. Functional architecture of macaque monkey visual cortex (Ferrier Lecture). Proc. R. Soc. Lond. B, 198, 1—59 (1977). Hubel D. H. Exploration of the primary visual cortex, 1955—78 (Nobel Lecture). Nature, 299, 512—524 (1982).
К главе 7.
Sperry, Roger. “Some effects of disconnecting the cerebral hemispheres” (Nobel Lecture, 8 Dec. 1981). In.: Les Prix Nobel, Almqvist & Wiksell International, Stockholm, 1982. Gazzaniga M. S., Bogen J. E., Sperry R. W. Observations on visual perception after disconnexion of the cerebral hemispheres in man. Brain, 88, 221—236 (1965). Gazzaniga M. S., Sperry R. W. Language after section of the cerebral commissures. Brain, 90, 131—148 (1967). Lepore F., Ptito M., Jasper H. H. Two Hemispheres — One Brain: Functions of the Corpus Callosum, Alan R. Liss, New York, 1984. Julesz, Bela. Foundations of Cyclopean Perception, University of Chicago Press, Chicago, 1971. Poggio G. F., Fischer B. Binocular interaction and depth sensitivity of striate and prestriate cortical neurons of the behaving rhesus monkey. J. Neurophysiol., 40, 1392—1405 (1977).
К главе 8.
Daw N. W. The psychology and physiology of colour vision. Trends in Neurosci, 7, 330—335 (1984). Hering, Ewald. Outlines of a Theory of the Light Sense (translated by Leo M. Hurvich and Dorothea Jameson), Harvard University Press, Cambridge, Mass., 1964. Ingle D. The goldfish as a Retinex animal. Science, 227, 651—654 (1985). Land E. H. An alternative technique for the computation of the designator in the Retinex theory of color vision. Proc. Natl. Acad. Sci. USA, 83, 3078—3080 (1986). Livingstone M. S., Hubel D. H. Anatomy and physiology of a color system in the primate visual cortex. J. Neurosci., 4, 309—356 (1984). Schnapf J. L., Baylor D. A. How photoreceptor cell respond to light. Sci. Am., 256, 40—47 (1987). Southall J.P.C. (ed.). Helmholtz’s Treatise on Physiological Optics (translated from 3d German edition), 3 vols. bound as 2, Dover Publishers, New York, 1962.
К главе 9.
Hubel D. H. Effects of deprivation on the visual cortex of cat and monkey, Harvey Lectures, Series 72, Academic Press, New York, 1978, pp. 1—51. Wiesel T. N. Postnatal development of the visual cortex and the influence of environment (Nobel Lecture). Nature, 299, 583—591 (1982). Wiesel T. N., Hubel D. H. Effects of visual deprivation on morphology and physiology of cells in the cat’s lateral geniculate body. J. Neurophysiol., 26, 978—993 (1963). Wiesel T. N., Hubel D. H. Receptive fields of cells in striate cortex of very young, visually inexperinced kittens. j. Neurophysiol., 26, 994—1002 (1963). Wiesel T. N., Hubel D. H. Single-cell responses in striate cortex of kittens deprived of vision in one eye. J. Neurophysiol., 26, 1003—1017 (1963). Wiesel T. N., Hubel D. H. Comparison of the effects of unilateral and bilateral eye closure on cortical unit responses in kittens. J. Neurophysiol., 28, 1029—1040 (1965). Wiesel T. N., Hubel D. H. Binocular interaction in striate cortex of kittens reared with artificial squint. J. Neurophysiol., 28, 1041—1059 (1965). Wiesel T. N., Hubel D. H. Extent of recovery from the effects of visual deprivation in kittens. J. Neurophysiol., 28, 1060—1072 (1965). Hubel D. H., Wiesel T. N., Le Vay S. Plasticity of ocular dominance columns in monkey striate cortex. Phil. Trans. R. Soc. Lond. B, 278, 377—409 (1977).
К главе 10.
Crick F.H.C. Thinking about the brain. Sci. Am., 241, 219—233 (1979). Hubel D. H. “Neurobiology: A science in need of a Copernicus,” in J. Neyman (ed.), The Heritage of Copernicus, Part II, M.I.T. Press, Cambridge, Mass., pp. 243—260. Van Essen D. C., Maunsell J. H. R. Hierarchical organization and functional streams in the visual cortex. Trends in Neurosci., 6, 370—375 (1983).
Последние комментарии
4 часов 1 минута назад
5 часов 48 минут назад
7 часов 2 минут назад
8 часов 7 минут назад
9 часов 16 минут назад
21 часов 14 минут назад